

Проводящие пути головного и спинного мозга
.pdf-20-
Пройдя через корешковую зону (zona radicularis) в боковой части заднего канатика, волокна входят в серое вещество заднего рога и заканчиваются на клетках грудного ядра (nucleus thoracicus), которые являются телами вторых нейронов данного пути.
Центральные отростки тел вторых нейронов выходят из серого вещества и направляются в боковой канатик спинного мозга на своей стороне. Совокупность аксонов вторых нейронов формирует восходящий пучок нервных волокон, который располагается в заднем отделе бокового канатика. На уровне поясничного отдела спинного мозга волокна от нейронов грудного ядра располагаются вблизи от задних корешков. Волокна из вышележащих сегментов примыкают к ним с вентральной стороны. Толщина пучка нарастает до уровня VII шейного сегмента спинного мозга. Далее волокна поднимаются в продолговатый мозг и через нижние ножки мозжечка проникают в мозжечок - к коре червя. Большая часть волокон в коре червя перекрещивается.
Волокна заднего спинно-мозжечкового пути в области спинного и продолговатого мозга не перекрещиваются, поэтому этот путь называют прямым или не перекрещенным мозжечковым путем.
Проводящие пути интероцептивного анализатора
От рецепторов внутренних органов поток импульсов направляется к коре головного мозга.
Все внутренние органы (относящиеся к пищеварительной, дыхательной, мочеполовой системам), а также стенки кровеносных сосудов содержат разнообразные чувствительные нервные окончания, которые называются интерорецепторы. Некоторые интерорецепторы в норме не вызывают ощущений. Другие интерорецепторы дают возможность ощутить жажду, голод, позывы на мочеиспускание и так далее. Рецепторы весьма многообразны и несут различную функциональную нагрузку. В зависимости от этого выделяют хеморецепторы - воспринимающие химические раздражения; механорецепторы - воспринимающие степень механического растяжения стенки сосудов, внутренних органов; терморецепторы.
Проводящие пути анализатора внутренних органов проходят в составе V, VII, IX, X черепных нервов, или в составе задних корешков спинного мозга.
От внутренних органов головы, шеи, грудной полости и частично от органов брюшной полости афферентные пути проходят в составе указан-
- 2 1 -
ных черепных нервов. Тело первого нейрона (псевдоуниполярные клетки) помещаются в одном из узлов черепных нервов. Периферические отростки в составе соответствующих нервов подходят к внутренним органам. Центральные отростки в составе корешков V, VII, IX, X черепных нервов входят в стволовой отдел мозга. Далее волокна этого пути подходят к нейронам следующих ядер ствола мозга, которые являются телами вторых нейронов:
1)вегетативные ядра соответствующих черепных нервов
2)ядро одиночного пути (nucleus tractus solitarius)
3)дорсальное висцеральное ядро (nucleus visceralis dorsalis) - располагается дорсальнее и медиальнее ядра одиночного пути
4)вставочное ядро - ядро Стадерини* (nucleus intercalatus) - располагается между дорсальным ядром блуждающего нерва и ядром подъязычного нерва.
Аксоны вторых нейронов переходят на противоположную сторону и в составе медиальной петли достигают таламуса. Тела третьих нейронов представлены клетками ядер зрительного бугра. Их аксоны формируют пучок волокон, который проходит через заднюю треть задней ножки внутренней капсулы и заканчивается на нейронах коркового ядра анализатора внутренних органов - в нижнем отделе постцентральной и предцентральнойизвилин.
Часть афферентных путей от органов брюшной полости и афферентные пути от органов малого таза идут в составе большого и малого чревных нервов (nervi splanchnici major et minor) и внутренних тазовых нервов (nervi splanchnici pelvini).
Тела первых нейронов располагаются в спинномозговых узлах (псевдоуниполярные клетки). Их аксоны в составе задних корешков входят в спинной мозг и заканчиваются на телах вторых нейронов - в промежуточной зоне серого вещества спинного мозга.
Аксоны тел вторых нейронов проходят в задних и боковых канатиках; в основном они присоединяются к tractus spinothalamicus и достигают ядер зрительного бугра, где расположены тела третьих нейронов. Их нейриты подходят к корковому ядру анализатора внутренних органов.
Афферентная иннервация внутренних органов не носит сегментарного характера; она имеет множество путей чувствительной иннервации. Основные пути иннервации идут из ближайших сегментов спинного моз-
* Стадерини (Staderini)- итальянский анатом, род. в 1869 г.

-22-
га. Дополнительные пути иннервации внутренних органов осуществляются из отдаленных сегментов спинного мозга. Таким образом, информация от внутренних органов поступает не только в ближайшие, но и в отдаленные сегменты спинного мозга. При повреждении спинного мозга дополнительные пути являются единственными афферентными путями от внутренних органов.
ЭФФЕРЕНТНЫЕ ПРОВОДЯЩИЕ ПУТИ
Эфферентные пути проводят нервные импульсы от различных центров головного мозга к рабочему органу - к двигательному и секреторному аппаратам.
Эфферентные пути делятся на две группы. Первую группу составляют пирамидные пути, начинающиеся пирамидными клетками в коре полушарий большого мозга. Ко второй группе относятся экстрапирамидные (внепирамидные) пути, проводящие импульсы от базальных ядер.
Эти пути двухнейронные. Тела первых нейронов располагаются в коре полушарий большого мозга (пирамидные пути), коре мозжечка, подкорковых ядрах (экстрапирамидная система). Мозжечок связан со спинным мозгом посредством промежуточных ядер ствола мозга (красное ядро, ядро оливы, ядра ретикулярной формации, латеральное преддверное ядро). Таким образом, импульсы от коры мозжечка поступают к клеткам указанных ядер, а далее следуют в нисходящем направлении к двигательным нейронам передних рогов спинного мозга. Сюда же подходят волокна всех нисходящих путей, следуя в боковых и передних канатиках спинного мозга.
Телами вторых нейронов эфферентных путей являются клетки двигательных ядер передних рогов спинного мозга, а также нейроны двигательных и секреторных ядер черепных нервов. Поток нисходящих импульсов усиливает или тормозит активность периферических (двигательных) нейронов, что ведет к изменению мышечного тонуса и двигательной активности мышц. Аксоны двигательных нейронов передних рогов спинного мозга выходят из спинного мозга в составе его передних корешков, далее в составе спинномозговых нервов и их ветвей в составе их ветвей достигают скелетной мускулатуры.
К двигательным нейронам передних рогов спинного мозга подходят волокна следующих эфферентных путей:
1) переднего корково-спинномозгового (пирамидного) пути;
-2 3 -
2)латерального корково-спинномозгового (пирамидного) пути;
3)красноядерно-спинномозгового пути;
4)покрышечно-спинномозгового пути;
5)оливо-спинномозгового пути.
В процессе филогенеза сначала сформировались эфферентные пути, проводящие импульсы из мозжечка. Они являются наиболее древними. Позднее развились пути, начинающиеся в покрышке среднего мозга. Наиболее молодыми являются пирамидные пути, которые формируются в связи с развитием новой коры полушарий большого мозга.
Пирамидные пути
Пирамидные пути осуществляют сознательное управление скелетной мускулатурой; служат для выполнения высокодифференцированных движений.
Наивысшего развития пирамидные пути достигли у человека. Это наиболее молодые эфферентные пути.
Волокна пирамидных путей направляются от пирамидных клеток коры полушарий большого мозга к нейронам двигательных ядер черепных нервов, либо передних рогов спинного мозга. В зависимости от этого пирамидные пути делят на корково-спинномозговые пути (передний и латеральный) и корково-ядерный путь. И те и другие пути начинаются большими пирамидными клетками, включая и гигантские пирамидные клетки Беца*, тела которых находятся в пятом слое коры предцентральной извилины полушарий большого мозга. По данным некоторых исследователей пирамидные пути начинаются также в коре постцентральной извилины, в задней трети верхней и средней лобных извилин, передней трети верхней теменной дольки и надкраевой извилины полушария большого мозга.
Пирамидные пути характеризуются некоторыми общими чертами, а именно:
-первыми нейронами путей являются большие пирамидные клетки, (в том числе и гигантские пирамидные клетки Беца), тела которых находятся в V слое коры полушарий большого мозга;
-в области полушарий большого мозга пирамидные пути следуют в составе лучистого венца и далее внутренней капсулы, занимая колено и передние две трети задней ножки последней;
*Бец В.А. - русский анатом, профессор Киевского университета (1834-1894)
-24- |
-25- |
-в стволе мозга пирамидные пути проходят последовательно через ножки мозга, мост, продолговатый мозг, располагаясь вентрально;
-на границе продолговатого и спинного мозга 80% волокон корко- во-спинномозгового пути переходит на противоположную сторону, образуя перекрест пирамид;
-в спинном мозге пирамидные пути следуют в передних канатиках (передний корково-спинномозговой путь) и в боковых канатиках спинного мозга (латеральный корково-спинномозговой путь);
-вторые нейроны пирамидных путей представлены клетками двигательных ядер черепных нервов (для корково-ядерного пути) или клетками двигательных ядер передних рогов серого вещества спинного мозга (для корково-спинномозгового пути);
-аксоны тел вторых нейронов в составе черепных или спинномозговых нервов достигают скелетной мускулатуры, где заканчиваются эффекторами.
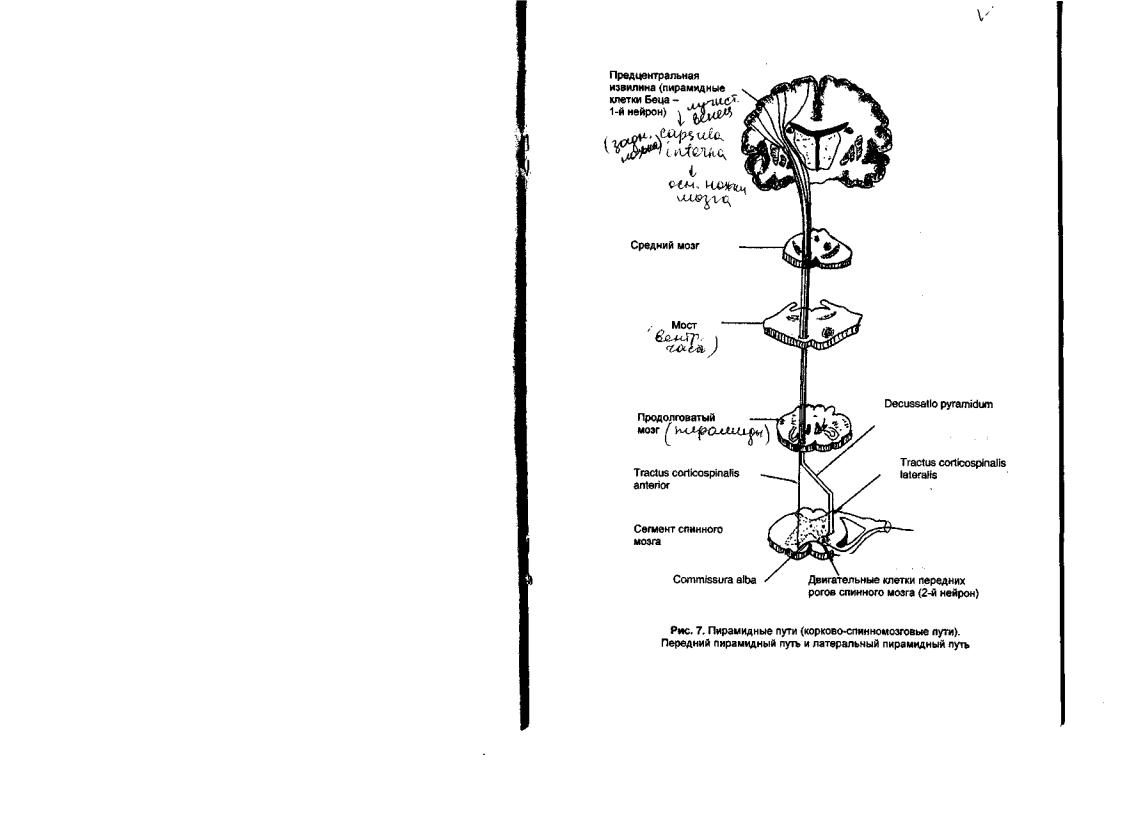
Корково-спинномозговойпуть-Tractuscorticospinalis
Корково-спинномозговой путь (рис. 7) проводит нервные импульсы от пирамидных клеток V слоя коры полушарий большого мозга к нейронам двигательных ядер передних рогов серого вещества спинного мозга.
Аксоны первых нейронов пирамидного пути идут в составе лучистого венца, а затем направляются между таламусом и чечевицеобразным ядром к внутренней капсуле, где проходят компактно, занимая передние две трети задней ножки внутренней капсулы. Во внутренней капсуле ближе к ее колену располагаются волокна, проводящие импульсы к мышцам верхней конечности, кзади от них проходят волокна для мышц туловища,
идалее — волокна для мышц нижней конечности. Затем волокна пути следуют через середину основания ножек мозга в вентральную часть моста. В области моста волокна корково-спинномозгового пути утрачивают компактность. Большое количество мелких пучков корково-спинномозгового пути отделяются друг от друга собственными ядрами моста и волокнами мостомозжечкового пути.
Впродолговатом мозге разрозненные пучки вновь собираются вместе
иформируют пирамиды (pyramis), которые выступают на вентральной поверхности мозга в виде продольных валиков. Итак, пирамиды, хорошо видимые на вентральной поверхности продолговатого мозга, представляют собой скопления волокон корково-спинномозгового пути.

-26-
На границе со спинным мозгом большая часть (80%) волокон пирамидного пути переходит на противоположную сторону, образуя пирамидный перекрест (decussatio pyramidum). Это "двигательный" или нижний перекрест в отличие от "чувствительного" - верхнего, образованного волокнами медиальных петель. Волокна корково-спинномозгового пути, которые перешли на противоположную сторону, проходят в боковых канатиках белого вещества спинного мозга и образуют латеральный корко- во-спинномозговой путь (tractus corticospinalis lateralis). Меньшая часть (20%) волокон остается на своей стороне, спускается в передних канатиках белого вещества спинного мозга, образуя передний корковоспинномозговой путь (tractus corticospinalis anterior).
Латеральный корково-спшшомозговой путь занимает внутреннезадний отдел бокового канатика и спускается в спинном мозге до его крестцового отдела. От поверхности спинного мозга этот путь отделяется слоем волокон заднего спинно-мозжечкового пути. По мере следования в каудальном направлении, от этого тракта отходят волокна, которые заканчиваются на клетках двигательных ядер передних рогов каждого сегмента. Значительно большее количество волокон отделяется от тракта в области утолщений спинного мозга, где в сегментах последнего располагается большее количество эффекторных нейронов, необходимое для иннервации массы мышц верхних и нижних конечностей. В боковом корковоспинномозговом пути наиболее медиальное положение занимают волокна, которые заканчиваются на нейронах шейных и грудных сегментов. Более латерально располагаются волокна спускающиеся до крестцовых и копчикового сегментов, то есть наиболее длинные волокна тракта. Передний корково-спинномозговой путь спускается в переднем канатике белого вещества спинного мозга, отделенный от передней срединной щели слоем волокон покрышечно-спинномозгового пути. В области передней белой спайки волокна переднего корково-спинномозгового пути посегментарно переходят на противоположную сторону и заканчиваются на двигательных нейронах передних рогов серого вещества спинного мозга. Передний корково-спинномозговой путь представляет собой небольшой пучок волокон, волокна его не опускаются ниже грудных сегментов спинного мозга.
Итак волокна переднего и латерального корково-спинномозговых путей переходят на противоположную сторону. Известно, что только 20% этих волокон заканчиваются непосредственно на двигательных нейронах передних рогов серого вещества спинного мозга, а 80% волокон подходит к вставочным нейронам, расположенным в передних рогах. Таким обра-
- 2 7 -
зом, двигательные нейроны передних рогов серого вещества спинного мозга могут являться третьими нейронами корково-спинномозговых путей.
На своем пути волокна пирамидных трактов отдают множество коллатералей, поэтому импульсы от каждого волокна направляются к нейронам нескольких сегментов одновременно. Установлено, что 55% всех волокон пирамидного пути заканчивается в шейных сегментах спинного мозга, 20% - в грудных, 25% - в поясничных сегментах. У взрослого человека 30% площади поперечного сечения спинного мозга приходится на пирамидный пучок.
Аксоны вторых нейронов (двигательных клеток передних рогов серого вещества спинного мозга) покидают спинной мозг в составе передних корешков; идут в составе спинномозговых нервов, их ветвей и достигают скелетной мускулатуры шеи, туловища, конечностей.
Выделяют три типа двигательных нейронов передних рогов серого вещества спинного мозга:
1. Большие альфа-клетки с толстым аксоном. Они вызывают быстрое сокращение мышц, потому что такой аксон проводит импульс с большой скоростью.
2.Малые альфа-клетки с тонким аксоном. Эти клетки поддерживают тонус мышц.
3.Гамма-клетки с очень тонким аксоном, который медленно проводит нервные импульсы. Большая часть движений начинается с активности гамма-клеток.
Существует строгая взаимосвязь между участками предцентральной извилины, парацентральной дольки (то есть корковым двигательным центром) и областью тела, то есть группой мышц, которой этот центр управляет. Иннервация мускулатуры является перекрестной, так как пирамидные пути перекрещиваются. Правое полушарие отвечает за движения левой полвины тела, и наоборот. Верхний отдел предцентральной извилины и парацентральной дольки посылает импульсы к мышцам нижней конечности, средний отдел извилины - к мышцам туловища и верхней конечности, и нижний - к мышцам шеи и головы (лица, языка, глотки, гортани). Схема Пенфилда* ("моторный гомункулус") дает представление о расположении в коре нейронов, дающих начало эфферентному пути для иннервации определенной группы мышц (рис. 8).
* Пенфилд В. (Penfild W.) - американский невролог

- 2 8 - |
-29- |
|
Нижние конечности человека выполняют в основном однообразные автоматизированные движения. В отличие от них движения верхних конечностей более разнообразны и сложны. Причем, деятельность правой и левой руки не идентична. Поэтому в тонком строении соответствующих отделов пирамидных путей должны присутствовать различия. Величина соответствующих областей коркового центра зависит от функциональной нагрузки - сложности движений различных групп мышц. Чем сложнее и дифференцированнее движения, выполняемые мышцами, тем больше корковый центр, связанный с этими мышцами. Так, почти вся средняя треть преддентральной извилины контролирует мышцы кисти и пальцев. Количество волокон пирамидного пути, посылающих импульсы к мускулатуре верхней конечности значительно больше, чем к нижней конечности.
Пирамидный путь формируется в процессе развития организма. У плодов 5-6 месяцев уже имеется пирамидный путь в виде пластинчатого тяжа, идущего от коры. Наиболее развит дорсальный его отдел. После рождения происходит увеличение объема пирамидного пути.
Значительное усложнение пути, особенно его вентрального отдела, наблюдается уже у детей школьного возраста.
При поражении пирамидных путей возникает нарушение произвольных движений в конечностях: полное отсутствие движений называется параличом; неполная утрата движений (уменьшение их силы и объема) называется парезом.
Характер паралича зависит от уровня поражения пирамидных путей. При поражении тела первого нейрона и его отростков возникает так называемый центральный паралич. Последний характеризуется отсутствием произвольных движений в соответствующих очагу поражения группах мышц при повышенных рефлекторных движениях (то есть повышении глубоких рефлексов) в этих же мышцах. Тонус мышц повышается. Такие нарушения объясняются тем, что при повреждении пирамидных систем тормозящее (субординирующее) влияние коры головного мозга не доходит до сегментарного аппарата спинного мозга. А поскольку сегментарный аппарат спинного мозга не поврежден, то он сохраняет свою рефлекторную деятельность.
При поражении двигательных клеток передних рогов серого вещества спинного мозга, его передних корешков, спинномозговых нервов (то есть тел вторых нейронов и его аксонов) развивается периферический паралич. Этот тип паралича характеризуется отсутствием как произвольных, так и рефлекторных движений в соответствующих группах мышц (утратой

-30-
глубоких рефлексов). Кроме того, выявляется снижение тонуса мышц, их атрофия. Гибель мышечных волокон сопровождается замещением их жировой и соединительной тканью. Симптомы такого поражения становятся понятны, если вспомнить, что нарушения происходят в рефлекторной дуге спинного мозга. При перерыве последней рефлекторный акт невозможен, мышца утрачивает свойственный ей постоянный тонус. Нарушение трофики мышц возникает в результате разобщения с клеткой (гамма нейрон) переднего рога, откуда по нервному волокну к мышце притекают нервнотрофические импульсы, стимулирующие нормальный обмен в мышечной ткани.
Корково-ядерныйпуть(Tractuscorticonudean's)
Этот путь (рис. 9) служит для сознательного управления поперечнополосатыми мышцами головы и шеи, которые иннервируются черпными нервами (III, IV, V, VI, VII, IX, X, XI, XII парами).
Это двигательный, двухнейронный путь, относящийся к пирамидной системе.
Тела первых нейронов корково-ядерного пути лежат в V слое коры в нижней трети предцентральной извилины. Аксоны клеток проходят в составе лучистого венца, далее — через колено внутренней капсулы и ствол мозга, где располагаются вентрально.
Часть волокон корково-ядерного пути постепенно переходит на противоположную сторону - к двигательным ядрам черепных нервов: в среднем мозге - к глазодвигательному и блоковому; в мосту - к тройничному, отводящему, лицевому; в продолговатом мозге - к языкоглоточному, блуждающему, добавочному, подъязычному. Другая часть волокон заканчивается на клетках этих же ядер на своей стороне. Таким образом, двигательные ядра черепных нервов являются телами вторых нейронов корко- во-ядерного пути. Аксоны вторых нейронов в составе черепных нервов подходят к мышцам головы и шеи.
Одностороннее поражение корково-ядерного пути вызывает парез - ограничение произвольных движений и снижение сократительной силы мышц. Неполная утрата функции соответствующих мышц связана с тем, что вследствие частичного перекреста пути клетки двигательных ядер черепных нервов получают импульсы от корковых центров обоих полушарий.

- 3 2 -
При двустороннем поражении корково-ядерных путей возникает центральный паралич, то есть полная утрата функции заинтересованных мышц. Исключением является подъязычный нерв. Корково-ядерный путь его полностью переходит* на противоположную сторону, поэтому поражение этого пути вызывает паралич (т.е. полную утрату функции) мышц языка с противоположной стороны.
Экстрапирамидная система
К экстрапирамидной системе относятся: таламус, хвостатое ядро, чечевицеобразное ядро, ограда, люисово тело, черное вещество, красное ядро, ядра ретикулярной формации мозгового ствола, нижнее оливное ядро продолговатого мозга, мозжечок и волокна белого вещества, связывающие эти ядра.
Впервые экстрапирамидная система появляется у рыб, усложняется у земноводных и пресмыкающихся, обеспечивая движения. У млекопитающих появляется пирамидная система, осуществляющая выполнение более совершенных, высокодифференцированных движений. В онтогенезе - экстрапирамидная система созревает раньше пирамидной. В отличие от пирамидной системы она имеет значительно больше переключений. Экстрапирамидная система занимает подчиненное положение. Экстрапирамидная система отвечает за непроизвольные автоматические движения - без участия сознания; регулирует тонус мышц и находится под контролем коры полушарий большого мозга.
Поражение экстрапирамидной системы вызывает изменение двигательной активности и тонуса мышц. При поражении хвостатого ядра и скорлупы появляются непроизвольные резкие движения (хореический гиперкинез). Поражение бледного шара проявляется скованностью мышц; нарушается пластический тонус мышц и движения в суставах становятся толчкообразными, замедленными. При поражении люисова тела появляются размашистые движения в конечностях.
Ядра экстрапирамидной системы имеют многочисленные связи между собой, а также с корой полушарий большого мозга.
От образований экстрапирамидной системы волокна нисходящих путей подходят к двигательным клеткам передних рогов серого вещества спинного мозга и формируют следующие пути:
1)красноядерно-спинномозговой;
2)покрышечно-спинномозговой;
-3 3 -
3)ретикуло-спинномозговой;
4)преддверно-спинномозговой.
Красное ядро является основным координационным центром экстрапирамидной системы. Оно характеризуется многочисленными связями с подкорковыми образованиями (таламусом, стриопаллидарной системой, подбугорной областью), а также с корой полушарий большого мозга.
Красноядерно-спинномозговойпуть(Tractusrubrospinalis)
Красноядерно-спинномозговой путь (монаковский) является наиболее мощным из экстрапирамидных путей (рис. 10). Это двухнейронный эфферентный путь автоматического действия. Начинается от крупных мультиполярных нейронов красного ядра, расположенного в покрышке среднего мозга. Аксоны клеток сразу переходят на противоположную сторону, образуя вентральный мезенцефалитический перекрест Фореля*. Далее волокна проходят через ствол мозга и следуют в боковых канатиках спинного мозга, посегментарно заканчиваясь на двигательных клетках передних рогов серого вещества спинного мозга. Клетки этих двигательных ядер являются вторыми нейронами пути. Аксоны вторых нейронов выходят из спинного мозга в составе передних корешков, а затем в составе спинномозговых нервов и их ветвей подходят к скелетной мускулатуре.
Покрышечно-спинномозговойпуть(Tractustectospinalis)
Покрышечно-спинномозговой путь - двигательный двухнейронный (рис. И). В филогенетическом отношении это один из наиболее древних путей, занимает подчиненное положение по отношению к коре полушарий большого мозга. Однако, этот зрительно-слуховой рефлекторный тракт осуществляет важные бессознательные защитные реакции в ответ на неожиданные зрительные и слуховые раздражения. Это так называемый четверохолмный рефлекс или старт-рефлекс, который подготавливает организм к быстрой реакции на новое внезапное раздражение.
Тела первых нейронов покрышечно-спинномозгового пути располагаются в сером веществе верхних и нижних холмиков среднего мозга. Здесь находятся одни из подкорковых центров зрения и слуха. Аксоны первых нейронов переходят на противоположную сторону под водопро-
* Форель (A. Fore!) - швейцарский невропатолог (1848-1931)

- 3 5 -
- 3 4 -

-36-
водом, образуя дорсальный перекрест, покрышки - фонтановидный перекрест Мейнерта**.
Далее волокна тракта идут в дорсальной части моста и продолговатого мозга, а затем в переднем канатике спинного мозга, где они прилежат вплотную к передней срединной щели. Аксоны первых нейронов посегментарно заканчиваются на клетках двигательных ядер передних рогов спинного мозга, которые являются вторыми нейронами покрышечноспинномозгового пути. Отростки вторых нейронов покидают спинной мозг в составе передних корешков и направляются к поперечнополосатым мышцам туловища и конечностей.
Меньшая часть волокон от холмиков среднего мозга подходит к двигательным ядрам черепных нервов (V, VII, XI, XII пары) и образует по- крышечно-бульбарный путь - tractus tectobulbaris. Аксоны нейронов двигательных ядер черепных нервов (тела вторых нейронов) в составе ветвей черепных нервов подходят к мышцам головы и шеи.
Ретикуло-спинномозговойпуть(Tractusreticulospinalis)
Этот тракт регулирует выполнение движений, требующих одновременного участия многих групп поперечнополосатых мышц. Импульсы, проходящие по волокнам ретикулоспинномозгового пути, оказывают тормозящее или активирующее влияние на нейроны спинного мозга.
Тела первых нейронов помещаются в стволе головного мозга - это крупные мультиполярные нейроны ретикулярной формации. Их аксоны в продолговатом мозге формируют компактный пучок волокон, который спускается в спикной мозг, где занимает латеральную часть переднего канатика и посегментарно заканчивается на двигательных клетках передних рогов серого вещества спинного мозга. Вторыми нейронами пути являются нейроны ядер передних рогов спинного мозга. Аксоны вторых нейронов выходят из спинного мозга в составе передних корешков, а далее через спинномозговые нервы, их ветви достигают скелетной мускулатуры.
- 3 7 -
Преддверно-спинномозговойпуть(Tractusvestibulospinalis)
Этот путь начинается от клеток латерального вестибулярного ядра (Дейтерса ), расположенного в дорсальной части моста. Волокна тракта проходят через продолговатый мозг и далее следуют в спинном мозге, где помещаются на границе переднего и бокового канатиков. Телами вторых нейронов являются двигательные клетки передних рогов спинного мозга. Их аксоны в составе передних корешков спинного мозга, спинномозговых нервов, их ветвей подходят к скелетным мышцам. Преддверно-спин- номозговой путь осуществляет влияние раздражений вестибулярных рецепторов на поперечно-полосатые мышцы.
Оливо-спинномозговойпуть(Бехтерева-Гельвега**) Tractusolivospinalis
Этот путь проводит координационные импульсы от центра равновесия (нижнего оливного ядра) к двигательным клеткам передних рогов серого вещества спинного мозга.
Тела первых нейронов располагаются в нижнем оливном ядре продолговатого мозга. Аксоны первых нейронов формируют пучок волокон, который проходит в передне-наружном отделе бокового канатика спинного мозга. Волокна оливоспинномозгового пути посегментарно подходят к двигательным клеткам передних рогов, которые являются вторыми нейронами пути. Аксоны вторых нейронов в составе передних корешков покидают спинной мозг, а далее в составе спинномозговых нервов их ветвей подходят к поперечно-полосатым мышцам.
Оливоспинномозговой путь определяется на уровне четырех верхних шейных сегментов спинного мозга.
Нисходящие пути от коры большого мозга к мозжечку Tractuscorticopontocerebellaris
Кора полушарий большого мозга регулирует деятельность мозжечка посредством корково-мосто-мозжечковых путей.
Путь двухнейронный, перекрещивается в области моста (рис. 12).
|
* Дейтерс О. (O.F. Deiters) - немецкий анатом и гистолог (1834-1863). |
"Мейнерт (Т. Meynert) - австрийский невропатолог (1833-1892) |
** Бехтерев В.М. - русский невропатолог, психиатр (1857-1927); Гельвег Г. (Helweg H) - дат- |
ский психиатр и физиолог (1847-1901). |

- 3 8 - |
-39- |
|
Тела первых нейронов помещаются в V слое коры полушарий большого мозга в лобной, теменной, височной и затылочной долях. Аксоны тел первых нейронов подходят к собственным ядрам моста на своей стороне, образуя корково-мостовой путь.
Взависимости от локализации первых нейронов, выделяют 4 пути:
1.Лобно-мостовой путь (tr. frontopontinus). Тела первых нейронов располагаются в коре лобной доли. Их аксоны формируют нервные волокна, которые проходят через задний отдел передней ножки внутренней капсулы и заканчиваются на клетках собственных ядер моста.
2.Теменно-мостовой путь (tr. parietopontinus). Тела первых нейронов
-в коре теменной доли.
3.Височно-мостовой путь (tr. temporopontinus). Тела первых нейронов помещаются в коре височной доли.
4.Затылочно-мостовой путь (tr. occipitopontinus). Тела первых нейронов локализуются в коре затылочной доли.
Волокна височно-мостового и затылочно-мостового пути образуют пучок, который проходит через среднюю часть задней ножки внутренней капсулы, основание ножки мозга и заканчивается на клетках собственных ядер моста. Это тела вторых нейронов корково-мосто-мозжечковых путей. Их аксоны, переходя на противоположную сторону, образуют мощный мосто-мозжечковый путь, который в составе средних ножек мозжечка достигает коры полушария мозжечка.
Эти нисходящие пути связывают кору каждого полушария большого мозга с корой полушарий мозжечка противоположной стороны.
Нисходящие двигательные пути мозжечка
Мозжечок контролирует деятельность двигательных нейронов передних рогов серого вещества спинного мозга, в том числе: поддержание равновесия, сохранение мышечного тонуса, преодоление инерции и силы тяжести. Этот контроль осуществляется через нисходящие двигательные пути мозжечка (рис. 13).
За импульсом произвольных движений (который по пирамидным путям направляется от полушарий большого мозга), следует мозжечковый импульс, осуществляющий поправку к основному импульсу. Мозжечковый импульс, достигая спинного мозга, вызывает возбуждение мышцантагонистов, останавливая тем самым движения по инерции, вызванное сокращением мышц от воздействия импульсов пирамидного пути.