

19.2. Развитие полового аппарата губок и кишечнополостных
У губок половой аппарат отсутствует. Половые клетки возникают из археоцитов и могут локализоваться в любой части тела (рис. 163). Из одного археоцита возникает либо одна овогония, либо кучка сперматозоидов. Последние часто бывают окружены особой питательной клеткой. У части губок несколько клеток образуют нечто вроде фолликула вокруг каждой овогонии, которая питается, фагоцитируя эти клетки. Сперматозоиды образуются преимущественно в наружных слоях паренхимы, овогонии – поблизости от жгутиковых камер. Губки гермафродитны или раздельнополы. Зрелые сперматозоиды удаляются через выносящие каналы наружу и оттуда, частично, через приносящие каналы и жгутиковые камеры, попадают в паренхиму женских зрелых особей, где оплодотворяют имеющиеся там зрелые яйца. Оплодотворенные яйца развиваются на месте, и в конце концов наружу выводятся готовые личинки.
Таким образом, половые клетки губок рассеяны внутри тела весьма широко, а единственные специальные приспособления, их обслуживающие, сводятся к питательным клеткам. Основную роль в выведении половых продуктов и в обеспечении встречи гамет играет, во-первых, собственная подвижность половых продуктов и, во-вторых, ирригационный аппарат, обслуживающий, как мы видели, почти все связи губки с внешним миром.
Кишечнополостные, если говорить об особи, а не колонии, стоят выше губок только на одну ступень. У гидроидов мы находим клетки, сходные с археоцитами, блуждающие но студенистой прослойке между эктодермой и энтодермой. Эти клетки оседают в определенных местах тела, и, размножаясь, дают начало небольшим яичникам или семенникам. У гидроидов эти гонады помещаются в эктодерме. При созревании половых продуктов прикрывающая их эктодерма лопается, и они выходят наружу. У гидромедуз гонады располагаются всегда поблизости от гастроваскулярного аппарата: либо в стенках хобота (Anthomedusae, Narcomedusae), либо на субумбрелле, по ходу радиальных каналов.
|
Рис. 163. Диффузное распределение половых клеток у низших Metazoa А – овоциты в теле губки Sycandra: 1 – хоаноциты, образующие эпителий жгутиковой камеры; 2 – иглы; в паренхиме видны каналы ирригационной системы, крупные овоциты и мелкие клетки паренхимы; Б – продольный разрез через гидрокаулон Eudendrium racemosum (Leptolida, Athecata): 1 и 2 – странствующие овогонии в эктодерме и энтодерме; 3 – молодой овогоний; 4 – интерстициальная клетка в энтодерме (по Н. Mcrgner); В – Oxyposthia praedator (Turbellaria, Acoela), расположение ово-гоний и овоцитов: 1 – осязательные сенсиллы; 2 – лобные железы; 3 – статоцист; 4 – рот; 5 – овогонии; 6 – овоциты; 7 – мужское половое отверстие (по А.В. Иванову)
|
|
|


У Scyphozoa гонады развиваются в энтодерме, но эктодерма субумбредлярной стороны образует против каждой из них мешочек (субгенитальную ямку), который, по-видимому, служит для подведения морской воды, а тем самым и кислорода, в непосредственную близость гонад. Выводятся половые продукты разрывом энтодермы – в полость кишечника, а оттуда через рот – наружу. У Anthozoa гонады также образуются в энтодерме, в толще перегородок в радиальных камерах, и половые продукты (а иногда зрелые личинки) выводятся наружу через рот.
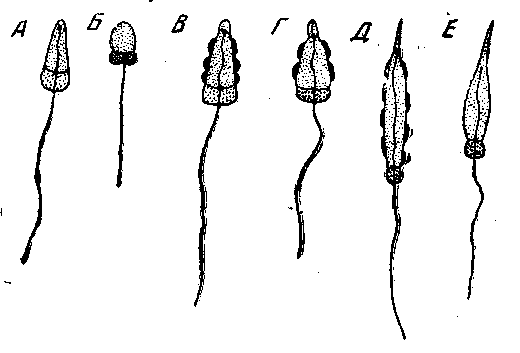
Рис. 164. Примитивные типы сперматозоидов Metazoa
А – Grantia compressa (Spongia, Calcarea); Б – Xenciurbella borfti Turbellaria, Xenoturbelllda); В – Tubularia meserribri/anthemum (Hydrozoa); Г – Paracentrotus lividus (Echinoidea); Д – Sfyela partite (Ascidia); E – Anguilla vulgaris (Pisces) (no O. Tuzet)
Сперматозоиды кишечнополостных очень близки по своему строению к таковым губок (рис. 164), что говорит против происхождения тех и других от различных групп Flagellata и в пользу того взгляда, что губки являются ранней боковой ветвью общего ствола Melazoa. Очень многие Bilateria имеют высокоспециализованные формы сперматозоидов, однако некоторые первичноротые, например одна из примитивнейших турбеллярий Xenoturbella bocki, а также многие низшие вторичноротые (иглокожие, асцидии) по строению сперматозоидов очень близки к кишечнополостным.
Выше мы видели, что Cnidaria, так же как и губки, почти лишены полового аппарата в тесном смысле этого слова. Но это относится только к особям. Колонии (по крайней мере колонии гидроидов), как правило, обладают колониальными половыми органами в виде гонофоров, несущих эти гонофоры бластостилей и т.п. Порой колониальный половой аппарат достигает значительной сложности – достаточно напомнить о корзиночках Plumulariidae (Thecaphora) и сходных образованиях. Более высокую органологическую дифференцировку колоний по сравнению с особями мы уже отмечали по поводу других аппаратов.
Гребневики, в отличие от большинства других кишечнополостных, являются гермафродитами. Половые продукты развиваются в энтодерме меридиональных каналов. Как принято думать, и яйца, и сперма выводятся наружу через рот. Однако A. Totton утверждает, что у большинства гребневиков яйца выводятся из меридиональных каналов через разрыв прикрывающей их эктодермы, а не через рот. У платиктенид семенники получают самостоятельные, открывающиеся непосредственно наружу семепроводы; у Coeloplana bocki T. Komai описывает тонкие канальцы, открывающиеся наружу 8-ью рядами отверстий и заканчивающиеся по направлению к яичникам пузырьками, иногда содержащими сперму. По-видимому, это семеприемники, содержащие сперму другой особи. Таким образом, у ползающих гребневиков мы впервые встречаемся с совокупительными органами, правда, еще очень несовершенными.