

18.3. Кровеносная система аннелид
Для типичных аннелид характерна высокая степень поляризации тканей: их тело построено из эпителиев, мышечной и нервной ткани. Все образованные этими тканями элементарные органы склеены между собой сравнительно тонкими пограничными пластинками. Плотное основное вещество этих пластинок почти лишено клеточных элементов. Пограничные образования существуют всюду, где сталкиваются два элементарных органа, например, два листка целомического эпителия. Все пограничные пластинки тела оказываются соединены между собой и образуют единую систему пограничных образований. Кровеносные сосуды представляют, как мы видели, систему каналов, возникающих в толще мезенхимы. У аннелид эта последняя оказалась зажата между стенками целома и прочих органов и превращена в систему пограничных образований. Вместе с ней оказалась зажата и кровеносная система, залегающая у аннелид в толще пограничных образований и пронизывающая эти последние.
Стенки сосудов аннелид образованы просто веществом пограничных пластинок. Мускулатура сосудов образуется за счет выселяющихся в рет-роперитонеальном направлении клеток целомического эпителия. Клапаны сосудов и так называемые сердечные тела (также служащие клапанами) образуются врастанием в полость сосуда того же целомического эпителия. Основной функцией кровеносного аппарата полихет является трофичеекая, как это видно из тесной связи этого аппарата с кишечником; кровь их богата растворенными белками и сравнительно бедна клеточными элементами. Содержание белков и глюкозы в крови наземных олигохет вполне сравнимо с содержанием их в крови человека. Наряду с этим кровь аннелид может играть роль и в газовом обмене; нередко она содержит в растворенном виде кровяные пигменты, гемоглобин или хлорокруорин, но почти никогда в ней не бывает окрашенных клеточных элементов. Зато она содержит неокрашенных фагоцитов. Как показал А.А. Заварзин для дождевого червя, малые фагоциты крови являются камбиальными элементами, за счет которых возобновляются в течение жизни животного все другие типы амебоцитов, а также и все клеточные элементы (фибробласты и т.д.), населяющие опорную ткань (пограничные образования). Этим подтверждается взгляд И.И. Мечникова о первичности трофической функции периферического фагоцитобласта и вторичности – опорной.
Первоначальной функцией дыхательных пигментов полихет является не перенос кислорода от дыхательных эпителиев к тканям, а накопление запаса кислорода на время периодических остановок дыхательных движений. В связи с этим в примитивнейших семействах полихет гемоглобин содержится не в крови, а в нервной ткани, наиболее чувствительной к недостатку кислорода. Гемоглобин в нервной ткани встречается и у немертин. В вышестоящих семействах полихет, например у Nereidae, гемоглобин содержится уже в крови, но основной его функцией продолжает оставаться создание переходящих запасов кислорода. Нахождение дыхательных пигментов в крови легко ведет к расширению их функций в сторону участия в разносе кислорода, что и становится в дальнейшем их основным назначением.
Наряду с примитивной структурой стенок сосудов кровеносный аппарат типичных аннелид обладает сложным архитектоническим расчленением (рис. 150). Наиболее постоянным компонентом кровеносного аппарата аннелид является околокишечный синус – узкое щелевидное пространство, заключенное в толще пограничной пластинки кишечника, между кишечным эпителием и спланхноплеврой. Оба листка пограничной пластинки, ограничивающие синус, бывают соединены между собой отдельными трабекулами основного вещества, пронизывающими полость синуса. У крупных аннелид, вместо сплошного околокишечного синуса, часто бывает околокишечное сплетение сосудов, точно так же муфтой одевающее кишечник. Околокишечный синус, или околокишечное сплетение бывают развиты только в постларвальных сегментах, не продолжаясь в ларвальные. Вместо этого, от спинной части околокишечного синуса отходит кпереди спинной сосуд, который и продолжается вдоль ларвальных сегментов, залегая в спинном мезентерии. При его основании у сидячих полихет часто бывает расположено сердечное тело, представляющее цилиндрический вырост стенки сосуда, лежащий в его просвете и направленный кпереди. При сокращении стенок сосуда волны перистальтики проходят сзади наперед; сокращенные участки сосуда прижимаются к сердечному телу и этим не допускают обратного тока крови кзади. Таким образом, сердечное тело функционирует в качестве клапана, лежащего при отхождении спинного сосуда от синуса.
Наряду с околокишечным синусом в брюшном мезентерии большинства аннелид залегает самостоятельный брюшной или подкишечный сосуд. У многих форм от околокишечного синуса на всем его протяжении обособляется и спинной сосуд. В этих случаях спинной и брюшной сосуды сливаются с синусом лишь в непосредственной близости заднего конца тела. Кроме того, сплетение сосудов или продольный сосуд обычно бывают и вдоль брюшной нервной цепочки. По околокишечному синусу и спинному сосуду кровь движется сзади наперед; по брюшному сосуду и периневральному сплетению – спереди назад. Эти продольные пути соединяются между собой метамерными сосудистыми дугами, первоначально расположенными в диссепиментах, т.е. интерсегментально. В состав каждой метамеры или ангиосомита входят две главные пары дуг – висцеральная и париетальная (рис. 151). Висцеральные дуги ведут кровь от спинного сосуда к стенкам кишечника, а от этих последних насыщенную питательными веществами кровь ведут к брюшному сосуду. Париетальные дуги ведут кровь от брюшного и околонервного сосудов к стенке тела и жабрам, где кровеносные сосуды распадаются на капилляры. В коже (рис. 151, В) и жабрах кровь насыщается кислородом.
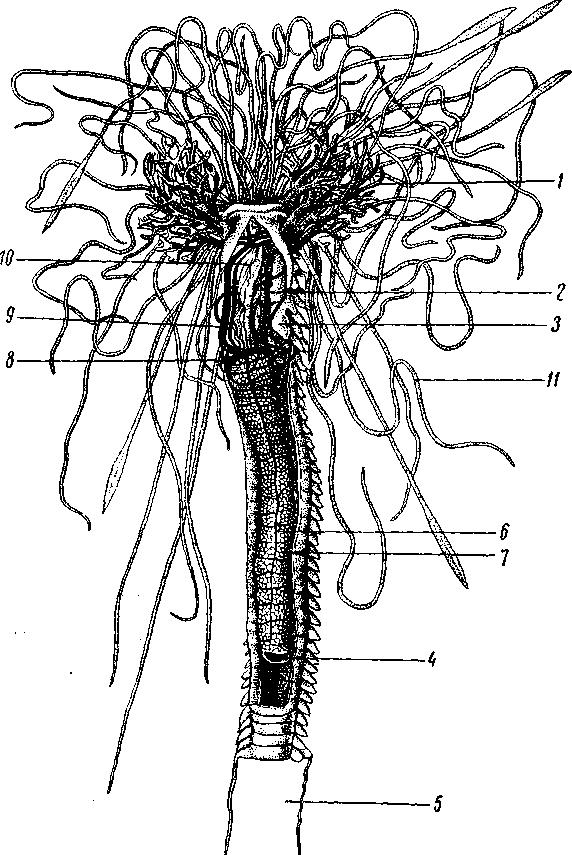
Рис. 150. Кровеносный аппарат Polymnia nebulosa (Polychaeta Terebellomorpha). Передняя часть тела вскрыта
1 – жабры; 2 – брюшной сосуд; 3 – нефромиксии; 4 – вентральный сосуд кишечника; 5 – жилая трубка; 6 – спинной сосуд на кишечнике; 7 – кишечник, сзади обрезанный; 8 – сосудистое кольцо вокруг пищевода; 9 – спинной сосуд; 10 – глотка; 11 – ловчие щупальца (по Cuvier)
Отсюда она возвращается отчасти к спинному сосуду, отчасти прямо к околокишечному синусу. Капилляры, проходящие в толще стенки тела, обеспечивают питание и газообмен мускулатуры и пр. Легко видеть, что при таком характере кровообращения в сегментах тела спинной сосуд содержит артериальную кровь, которую он и несет к надглоточному ганглию и органам чувств головы. У всех полихет значительные отличия представляет кровеносный аппарат передних сегментов тела благодаря сосудам, отходящим к глоточному аппарату. Кроме того, основной план ангиосомита подвергается бесчисленным видоизменениям, во-первых, в разных семействах полихет и олигохет и, во-вторых, при наличии гетерономное – в разных отделах тела и сегментах одного итого же животного. Основной путь развития кровеносного аппарата аннелид ведет к возрастанию гетерономности ангиомерии, обусловливающей возрастающую интеграцию и централизацию всего кровеносного аппарата в целом.
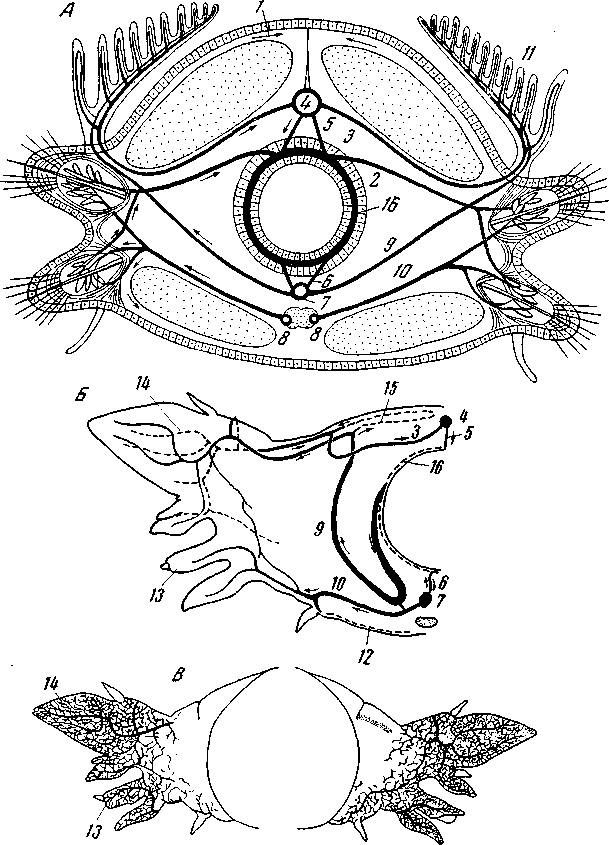
Рис. 151. Ангиосомит полихет
А – схематический поперечный разрез через туловищный сегмент снабженной жабрами блуждающей полихеты; сосуды показаны черным; мелкие сосуды в стенке тела и мускулатуре не изображены (по Догелю); Nereis virens: Б – ангиосомит; В – капиллярная сеть в коже параподиев (по Lindroth); 1 – париетальные сосуды; 2 – параподио-интестинальный; 3 – параподио-дорсальный; 4 – дорсальный; 5 – дорсо-интестинальный; 6 – интестино-вентральный; 7 – вентральный; 8 – невральные; 9 – вентропараподиальные; 10 – невропараподиальные; сосуды или их гомологи; 11 – жабра; сеть кожных капилляров: 12 – вентральная; 13 – невроподиальная; 14 – нотоподиальная; I5 – дорсальная; 16 – кишечный синус (или кишечное сплетение)
Однако в соответствии с чрезвычайным разнообразием гетерономной дифференцировки сегментов тела, которое мы наблюдаем у полихет, направления развития кровеносного аппарата также представляют крайнее разнообразие.
Итак, для аннелид характерна, с одной стороны, довольно низкая ступень гистологической дифференцировки кровеносного аппарата, с другой – полная замкнутость и далеко идущее архитектоническое расчленение этого аппарата. Правильная циркуляция крови существует, но пропульсаторная функция мало централизована. Единого сердца, обеспечивающего продвижение крови по сосудам, здесь нет. Кровь движется благодаря сократимости больших участков продольных сосудов, а часто и передних дуг. Ритмы отдельных сосудов согласованы очень несовершенно, и вообще работа всего кровеносного аппарата мало интегрирована. Артериальное давление Nereis в покое – всего на 1,0–1,5 мм рт. ст. выше давления целомической жидкости; при движении червя разность подымается до 6 мм.
О редукции кровеносного аппарата у ряда форм и функциональной замене его целомом было сказано выше.
Сравнивая кровеносный аппарат аннелид и немертин, мы находим значительное сходство в гистологической структуре и резкое различие в общем плане расположения сосудов. У немертин нет ничего подобного околокишечному синусу, составляющему основную и наиболее постоянную часть кровеносного аппарата полихет. Наряду с кишечным синусом, у аннелид сильно развиты непарные продольные сосуды – спинной, брюшной, околонервный, тогда как у немертин – парные боковые. У полихет парные продольные сосуды, если и встречаются, то лишь вторично, в наиболее специализованных типах кровеносного аппарата (например, у те-ребеллид), да и здесь лишь на небольшом протяжении тела. Коренное различие в плане строения кровеносного аппарата немертин и аннелид стоит в полном соответствии с представлением о независимом возникновении этого аппарата в обеих группах, в свою очередь вытекающим из обоснованного положения об отсутствии близкого родства между сколецидами и трохофорными животными.