

8.3. Метамерия туловища насекомых
Теперь перейдем к рассмотрению метамерии туловища насекомых. Характернейшей особенностью насекомых, в отличие от многоножек, является разбивка их туловища на два отдела, или тагмы: грудь и брюшко. Грудь всецело берет на себя локомоторную функцию: здесь сохраняются ноги (локомоторные конечности), и у высших насекомых (Pterygota) возникают крылья. Брюшко, наоборот, большей частью теряет все или почти все конечности; если же последние частично и сохраняются, они перестают активно участвовать в передвижении животного; они принимают на себя функцию половых придатков, органов чувств и т.п. или низводятся на степень простых рудиментов.
Исключение составляют главным образом брюшные ножки некоторых личинок, а во взрослом состоянии – вилка Collembola. Итак, грудь насекомого почти всецело берет на себя функцию передвижения. В силу этого мускулатура грудных сегментов усиливается, диаметр их увеличивается, наружный скелет осложняется. В сегментах брюшка, наоборот, мускулатура и скелет упрощаются; по толщине сегменты брюшка часто значительно уступают сегментам груди. У хорошо летающих насекомых (Diptera, Hymenoptera) происходит все возрастающая консолидация груди, интеграция ее сегментов в одно сложное целое и одновременная дифференцировка сегментов брюшка, ведущая к резко выраженной гетерономности этого последнего. Таким образом, от почти гомономного, удлиненного туловища наиболее примитивных насекомых, еще напоминающего туловище многоножек, мы видим ряд переходов к высокогетерономному и интегрированному туловищу высших насекомых с мощной локомоторной грудью и компактным дифференцированным брюшком. Насекомые в современном объеме этого класса вряд ли являются монофилетической группой. Из Apterygota только Thysanura, несомненно, близки крылатым насекомым, тогда как Campodeoidea, Protura и, в особенности, Collembola занимают весьма обособленное положение. Возможно, что все они являются самостоятельными группами, наподобие Symphyla или Chilopoda.
Campodeoidea (рис. 56, Б, В) живут, подобно Symphyla, в почве и лесной подстилке. Грудные сегменты уже и здесь несколько отличаются от брюшных большим диаметром, более сложным скелетом, присутствием ходильных ног. 1-й сегмент груди несколько меньше остальных. Все сегменты груди подвижно сочленены между собой. Сегментов брюшка 11; 11-й сегмент рудиментарный, но несет пару церков; 8–10-й – тоже несколько меньше остальных; 1-й сегмент брюшка несет пару двучленистых рудиментарных ножек; 2–7-й сегменты несут придатки в виде неподвижно приросших коксоподитов, к каждому из которых прикрепляются по 1 грифельку (stylus) и по 2 брюшных мешочка, напоминающих грифельки и брюшные мешочки Symphyla. Грифельки, по мнению различных авторов, соответствуют или экзоподитам двуразветвленной конечности, или, что более вероятно, эпиподитам. Конечности половых сегментов редуцированы. Представители отряда Thysanura (рис. 57) в некоторых отношениях построены еще гомономнее; в частности, они имеют грифельки на всех сегментах брюшка – со 2-го по 9-й включительно; на 8 и 9-м сегментах имеются, кроме того, и половые придатки – гонапофизы (рис. 58), представляющие неотчлененные выросты коксоподитов.
У Machilidae (рис. 56, Г) грифельки имеются и на коксах двух задних пар грудных ног. Сравнительно хорошо развитый 11-й сегмент брюшка несет церки и сверх того – спинную медиальную нить. Гомономность тела несколько нарушается его веретеновидной формой, очевидно связанной с возросшей подвижностью и открытым образом жизни чешуйниц. Однако личинка еще не веретеновидна и очень гомономна. Веретеновидная форма тела обусловлена аксиальной дифференцировкой диаметра сегментов и не связана с глубокой перестройкой самих сегментов.
Таким образом, необходимая для обеспечения быстрых движений интеграция тела является у Thysanura значительно более поверхностной, чем та интеграция, которую мы увидим у высших насекомых. Thysanura известны с триаса, родственные им Monura – с карбона.
Весьма гомономное строение и у представителей отряда Protura, имеющих три пары абдоминальных конечностей (рис. 56, А) – настоящих ножек. В семействе Eosentomidae все они двучленистые, у Acerentomidae по меньшей мере одна пара (третья) одночленистая. Особенности Protura, отличающие их от всех остальных насекомых, – сильное развитие анальной лопасти и расположение полового отверстия на заднем краю 11-го сегмента. Эти образования в некоторых группах Collembola могут подвергаться и редукции.
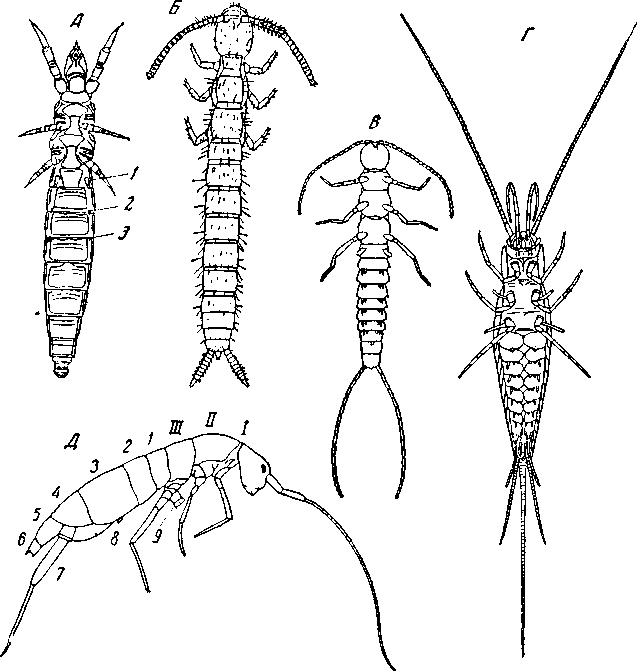
Рис. 56. Расчленение тела Insecta Apterygota
А – Acerentomon doderoi (Protura), с брюшной стороны: 1, 2,3 – ножки первых трех сегментов брюшка (по Berlese); Б – Anajapyx vesiculosus (Campodeoidea) (no Silvestri); 11-й членик брюшка рудиментарен, но несет церки; между ними видна анальная лопасть; В – Campodea staphylinus (Campodeoidea) (noTillyard); Г – Machilis sp. (Thysanura) (no Handlirsch); Д – Tamocerus plumbeus (Collembola) (no Willem): 1–6 – членики брюшка; 7 – вилка; 8 – зацепка; 9 – брюшная трубка; I, II, III – передне-, средне- и заднегрудь
Collembola (рис. 56, Д) имеют в строении и развитии ряд признаков, общих с Diplopoda и Symphyla и отсутствующих у других насекомых. Как мы видели выше, они имеют неполное число сегментов туловища, совпадающее с числом их у первой личинки Symphyla, до начала анаморфоза. Сегменты груди, как и у других Apterygota, не слиты между собою, однако они различной величины: 1-й из них обычно меньше всех, 2-й – больше всех. Сегменты брюшка также неравны между собой и несут 3 пары придатков: 1-й сегмент несет брюшную трубку (tubus ventralis), которая соответствует паре слившихся брюшных мешочков; 4-й сегмент несет прыгательную вилку (furca), образовавшуюся частичным слиянием пары конечностей; конечности 3-го сегмента иногда сохраняются в виде так называемой зацепки (hamulus).
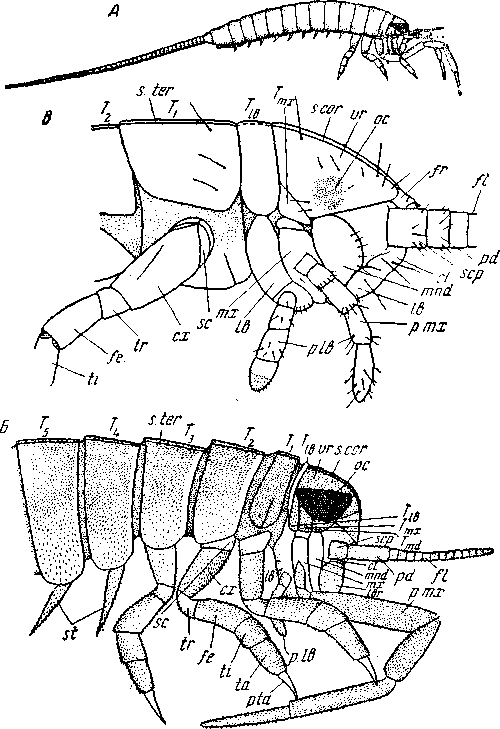
А – Dasyleptus brogniarti (Monura), реконструкция, вид слева; Б – то же, голова и передняя часть туловища; В – Lepisma saccharinum (Thy-sanura), голова и передняя часть туловища ларвулы I возраста. cl – наличник; сх – кокса; fe – бедро; fl – жгут антенны; fr – лобный склерит; lb – нижняя губа (максиллы вторые); lbr – верхняя губа; mnd – мандибулы; тх – максиллы первые; ос – боковые глава; pd – pedicellus (2-й членик антенны); р. тх – максиллярные пальпы; р. lb – нижнегубные пальпы; pta – praetarsus; sc – субкокса; s. cor – эпикраниальный шов; scp – scapus (1-й членик антенны); st – грифельки (styli); s. ter – тергальный шов; ta – лапка (tarsus); ti – голень; tr – вертлуг; Tmd, Ттх, Тlb, – тергиты мандибулярного, максиллярного и лабиального сегментов; T1-T5 – тергиты 1–5-го туловищных сегментов, vr – теменной склерит (по Д.Г. Шарову)
Ни гонапофиз, ни грифельков, ни церков у Collembola не бывает. Два последних сегмента брюшка значительно меньше остальных. Итак, мы видим, что с точки зрения дифференцировки своих туловищных сегментов Collembola также отличаются самобытностью. Они представляют в эволюции членистоногих ранний пример укорочения брюшка и дифференцировки его сегментов; процесс этот разыгрывался, очевидно, на такой ступени организации, когда брюшко еще было богато конечностями и брюшными мешочками, которые и получили своеобразное развитие в связи с возникновением у Collembola нового способа передвижения – прыгания при помощи вилки. Collembola отличаются очень большой древностью, так как вполне типичный их представитель – Rhyniella praecursor – известен из девона.
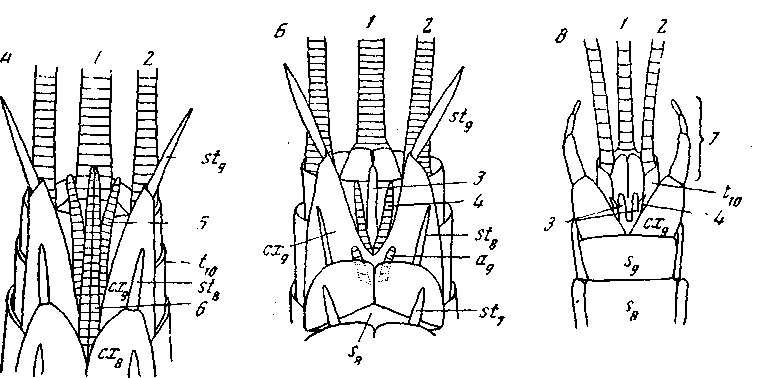
Рис. 58. Придатки заднего конца брюшка низших насекомых,
вид с вентральной стороны
А – Machilis (Thysanura), самка; Б – Machilis, самец; В – поденка (Ephemeroidea), самец; все – схематически: 1 – непарная хвостовая нить; 2 – церки; 3 – пенис (парный у поденок); 4 – парамер; створки яйцеклада: 5 – наружные; 6 – внутренние; 7 – щипцы (claspers) самца; сх8–сх9 – коксы 8-9-го сегментов, [sf7 – st9 – styli 7–9-го сегментов, s8 и s9 – стерниты 8–9-го сегментов; t10 – тергит 10-го сегмента (из Imms)