

2.7. Вопросы для самоконтроля
1. Какой симметрией обладают губки?
2. Какие виды симметрии встречаются у кишечнополостных, полипов и гидромедуз?
3. Симметрия сцифомедуз?
4. Особенности симметрии коралловых полипов.
5. Каково строение взрослого гребневика?
6. Особенности симметрии гребневиков.
7. Сравнение симметрии гребневиков и кишечнополостных.
8. Экологическое значение симметрии губок и кишечнополостных.
ТЕМА 3
АРХИТЕКТОНИКА ПЛОСКИХ ЧЕРВЕЙ
3.1. Симметрия турбеллярий
Среди первичноротых наиболее примитивной группой как по общему уровню развития, так, в частности, и по типу своей симметрии являются плоские черви (Plathelmintes) и в особенности турбеллярии (Turbellaria).
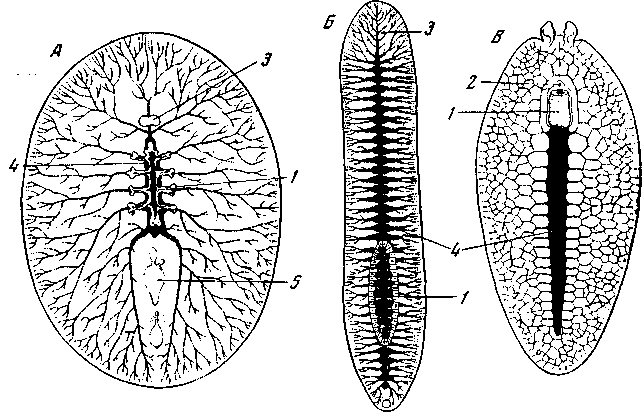
Рис. 16. Поликлады
Экологически двусторонняя симметрия турбеллярии явно связана со способом передвижения большинства представителей этого класса – с ползанием по дну, поведшим к дифференцировке спинной и брюшной сторон. Наоборот, правая и левая стороны ползающего животного сохраняют симметрическое подобие в связи с тем, что нарушение его вызвало бы неравномерное давление воды на обе половины тела движущегося животного и затруднило бы движение. Так же обычно объясняется возникновение двусторонней симметрии и у других Bilateria, к чему мы будем еще иметь случай вернуться.
Существуют две теории, выводящие симметрию турбеллярии из радиальной симметрии кишечнополостных. Одна из них принадлежит А. Лангу, другая – Л.Ф. Граффу.
A. Лaнг (1884) выводит плоских, широких поликлад, вроде Planocera и Stylochus, из сплющенных в направлении главной оси тела гребневиков отряда Platyctenida (Ctenoplana, Coeloplana) и приравнивает дорсовентральную ось поликлад главной оси тела гребневиков.
Хорошо разработанная и по тому времени хорошо обоснованная теория Ланга долгое время пользовалась большим успехом и являлась общепризнанной. Однако по существу эта теория основана главным образом на рассмотрении одной группы – поликлад и преимущественно строения их кишечного аппарата. Нервный аппарат поликлад Ланг толкует, как мы увидим, неправильно. Строение полового аппарата и других органов он вообще во внимание не принимает. Поэтому в настоящее время теория Ланга нас больше удовлетворять не может.
Л.Ф. Граф (1905) считает наиболее примитивными из турбеллярий Acoela, сравнивает их с личинками кишечнополостных и склонен производить их из гипотетических планулообразных предков этих последних. С точки зрения теории первичной седентарности кишечнополостных (А. Захваткин, 1949), Acoela произошли от каких-то древних кишечнополостных путем прогрессивной неотении, т.е. путем достижения половой зрелости на стадии паренхимулы, сохранения ею подвижного образа жизни и усовершенствования ее организации в соответствии с таким образом жизни.
Многие Acoela имеют продолговатое (в разрезе круглое) тело; все они, подобно плануле, покрыты с поверхности мерцательным эпителием; у юных особей рот может находиться почти на самом заднем конце тела. Внутри тело заполнено паренхимой, периферическая часть которой носит преимущественно опорный характер, а центральная – преимущественно пищеварительный. Передний конец тела Acoela, как и у планулы, богаче нервными элементами, чем задний. Вблизи переднего конца у всех Acoela имеется статоцист, нередко окруженный мозгом. У чрезвычайно примитивной Xenoturbella bocki (отряд Xenoturbellida, близкий к Acoela) статоцист лежит в эпителии переднего конца тела, приближаясь в этом отношении к аборальному органу гребневиков.
Графф проводит сравнение следующим образом: передний конец бескишечной турбеллярий он приравнивает аборальному концу планулы, задний конец – оральному. Главная ось планулы оказывается гомологичной продольной оси турбеллярий (а не дорсовентральной оси, как в теории Ланга). Фактически из всех известных нам бескишечных турбеллярий рот лежит точно на заднем конце тела только у Diopisthoporus (рис. 17, Б). Лучевая симметрия Diopisthoporus настолько хорошо выражена, что спинную и брюшную стороны у неполовозрелых или у женских зрелых особей различить бывает нелегко. Двусторонняя симметрия проявляется главным образом в строении мужского полового аппарата (рис. 17, В). Женскую особь Diopisthoporus можно считать животным с почти полной лучевой симметрией; подобно планулам кишечнополостных, она имеет ось вращения неопределенно большого порядка. Однако у огромного большинства взрослых Acoela рот смещается с заднего конца тела по направлению кпереди и этим лучевая симметрия нарушается (рис. 17, Г). После такого смещения остается одна единственная плоскость симметрии, проходящая через оба полюса и рот; она является сагиттальной плоскостью. Та сторона тела, на которой оказывается рот, этим самым становится брюшной стороной, противоположная – спинной. Так возникает, согласно теории Граффа, двусторонняя симметрия турбеллярий.
Рот поликлад бывает расположен очень различно (рис. 16), и наиболее примитивным является его расположение в задней части тела и отхождение глотки от заднего конца главной кишки, как, например, у Cestoplana. В пользу первичности положения рта на заднем конце тела говорит то обстоятельство, что у зародышей почти всех турбеллярий, а не только бескишечных, рот находится или на заднем конце тела, или вблизи него. Лишь на дальнейших стадиях онтогенеза происходит разрастание заднего конца и рот сдвигается вперед вдоль брюшной стороны. Но почему считать, что оральный полюс турбеллярий все еще находится на заднем конце тела, если рот больше не лежит на этом конце? Ведь именно присутствие рта и является, как правило, наиболее характерным признаком орального полюса. В пользу того, что оральный полюс тела сохраняет у бескишечных свое место на заднем конце тела, говорит прежде всего строение их нервного аппарата, подробно изученное самим L. v. Graff (рис.17. Д).
Acoela обычно имеют несколько пар продольных нервных стволов. Стволы эти идут меридионально, соединены многочисленными кольцевыми комиссурами и представляют по своему расположению более или менее совершенную лучевую симметрию.
Осью этой симметрии является прямая линия, соединяющая передний и задний концы тела. Кожная мускулатура бескишечных, образующая правильную решетку из кольцевых и продольных волокон, также обладает лучевой симметрией неопределенно многолучевого типа по отношению к той же продольной оси и совершенно независимой от положения рта. Таким образом, положение рта весьма мало отражается у турбеллярий на проявлении остаточной лучевой симметрии в их теле. Это позволяет нам сказать, что у этих животных происходит не искривление главной оси, а лишь смещение рта по отношению к другим системам органов, более или менее сохраняющим свою первоначальную симметрию. Это смещение, почти не нарушая первоначальную лучевую симметрию нервного аппарата или кожной мускулатуры, нарушает общую лучевую симметрию тела. Лучевая симметрия, которая после этого остается в строении нервного аппарата и других систем органов, обуславливает собой лишь неполную, остаточную лучевую симметрию тела турбеллярий, сохраняющуюся в нем наряду с господствующей двусторонней симметрией.
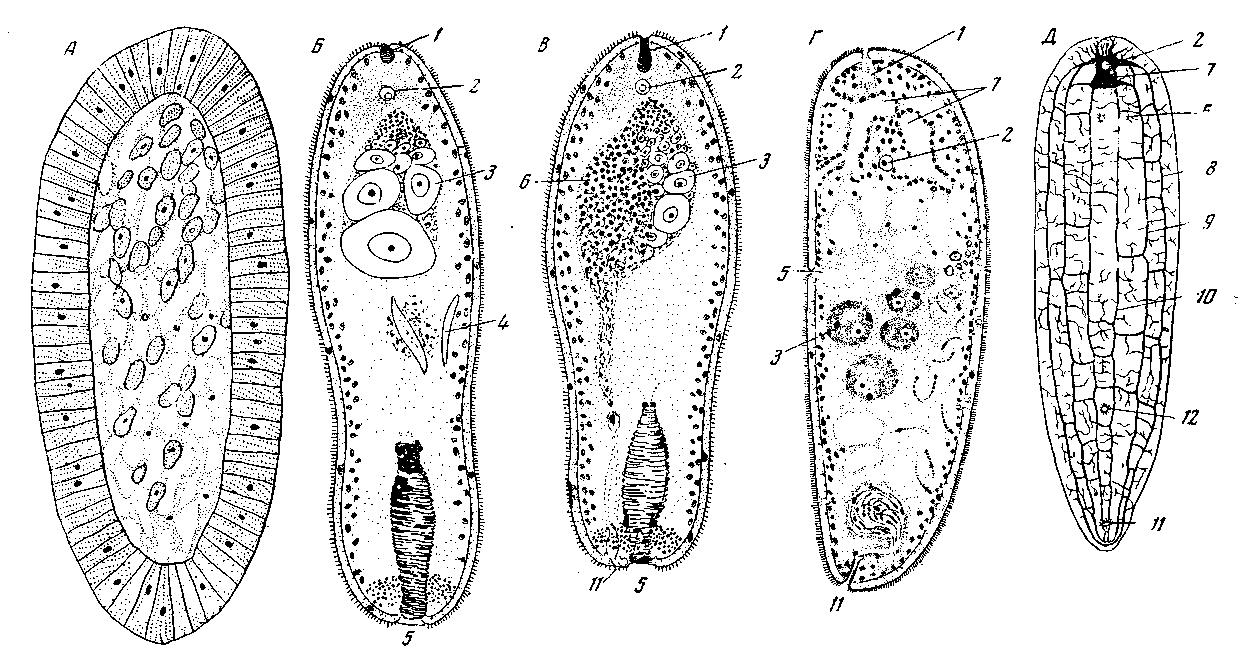
Рис. 17. Возникновение двусторонней симметрии турбеллярий из радиальной симметрии планулообразных форм, согласно взглядам L. v. Graff
А – планула гидромедузы Turritopsis; Б – Diopisthoporits longitubua (Turbellaria Acoela), женский экземпляр, вид с правой стороны; пример турбеллярий с почти ненарушенной радиальной симметрией; В – то же, гермафродитный экземпляр, благодаря положению мужского полового отверстия животное приобретает некоторую степень двусторонней симметрии (по Westblad); Г – Haploposthia Ъгиппеа (Acoela), вид с левой стороны, рот и половое отверстие смещены на одну из меридиональных сторон, которая в силу этого определяется как брюшная сторона, животное приобретает хорошо выраженную двустороннюю симметрию (по An der Lan); Д – Convolula rosholfensis, нервный аппарат, вид со спинной стороны, при господстве двусторонней симметрии остатки радиальности в расположении продольных нервных стволов (по Graff): 1 – лобные (теменные) железы; 2 – статоцист; 3 – яичники; 4 – пищевые частицы в паренхиме; 5 – рот; 6 – семенники; 7 – мозг; продольные нервные стволы: 8 – боковые; 9 – наружные брюшные; 10 – внутренние брюшные; половое отверстие: 11 – мужское; 12– женское