

Теории антителообразования и взаимодействия антител с антигенами
Теория «боковых цепей». Механизм образования антител и их взаимодействие с антигенами впервые обосновал в теории «боковых цепей» П. Эрлих (1898). Представив клетку в виде атомных групп и радикалов, он как химик полагал, что центрально расположенные группировки могут определять ее видовую специфичность, а поверхностные («боковые цепи») - выполнять функцию рецепторов антигенов. Соединяясь с ними, клетка, по мнению П. Эрлиха, затем вырабатывает их так много, что они, не удерживаясь на мембране, отщепляются и поступают в кровь, где функционируют как антитела, способные связывать микробы и токсины. При этом П. Эрлих представлял, что клетка якобы может иметь антитела трех порядков: антитела первого порядка, обусловливающие нейтрализацию токсина, имеют одну группу связывания, которая соединяется с гаптеном антигена; антитела второго порядка, вызывающие реакции агглютинации и преципитации, обладают группой связывания и эффекторной группой склеивания или осаждения антигенов; антитела третьего порядка, связывающие комплемент, содержат две группы связывания: одна вступает в реакцию с антигеном, а другая -с комплементом.
Современные теории антителообразования подразделяют на три группы: 1) инструктивные - теории прямой и непрямой матрицы; 2) селекционные - теории непрямого отбора и селекции клонов антителообразующих клеток; 3) дерепрессии генов.
Теория прямой матрицы. Выдвигая эту теорию в 1930 г., Ф. Брейнль и Ф. Гауровитц предполагали, что синтез антител в клетках происходит на детерминантах антигена, которые, являясь матрицей, определяют структуру и конфигурацию антигенсвязывающих центров (рис. 66, а). Выработанные таким образом под воздействием антигенов иммуноглобулины с центрами связывания в виде слепков детерминант антигена приобретают высокую степень специфичности и способность реагирования лишь с комплементарными гаптенами.
Теория непрямой матрицы. Создатели этой теории Ф. Вернет и Ф. Финнер в целом разделяя идею матричного характера формирования специфичности иммуноглобулинов, в 1949 г. высказали гипотезу опосредованного воздействия антигенов на генотип антителообразующих клеток (рис. 66, б). При этом непрямая информация образования комплементарных антигену иммуноглобулинов передается потомкам клеток.
 Большой
научной ценности теория «боковых цепей»
и матричные теории антителообразования
в настоящее время не имеют, но на их
основе формировались селекционые
теории и теория дерепрессии генов.
Большой
научной ценности теория «боковых цепей»
и матричные теории антителообразования
в настоящее время не имеют, но на их
основе формировались селекционые
теории и теория дерепрессии генов.
Теория непрямого отбора. Выдвинутая в 1955 г. Н. Йерне теория непрямого отбора допускала наличие в организме многочисленных антителопродуцирующих клеток, а в крови - спонтанно выработанного огромного набора антител ко всевозможным антигенам. Антителообразование к каждому из них объяснялось тем, что комплекс антитело - антиген побуждал выработку только таких иммуноглобулинов, которые в нем содержались. Таким образом, антиген в антител ообразовании выполнял роль селектора, а не сигнального индуктора.
Теория селекции клонов антителообразующих клеток. Разработанная Ф. Бернетом в 1959 г. теория селекции клонов антителообразующих клеток постулирует: в процессе длительной эволюции в организме сформировалась гетерогенная популяция антителообразующих клеток, способных продуцировать антитела определенной специфичности; антителообразование детерминировано генотипом и не зависит от антигена; антиген лишь стимулирует (селекционирует) клоны тех антителообразующих клеток, которые имеют к нему специфические рецепторы; в ответ на антигенный стимул происходит пролиферация антителообразующих клеток, а параллельно - и дифференциация предадаптированных клеток в плазмоциты, продуцирующие преддетерминированные по структуре и, главное, - специфичности - иммуноглобулины (рис. 67). Теория Ф. Бернета допускает, что в момент перехода исходных клеток в дифференциацию они могут вырабатывать несколько видов иммуноглобулинов. В дальнейшем в присутствии антигена-индуктора одно направление синтеза иммуноглобулина закрепляется, а остальные репрессируются. Те клетки, в которых синтез иммуноглобулина не закрепляется, погибают. Кроме того, в результате взаимодействия антигена образуются не до конца дифференцированные клетки иммунологической памяти. Возможно, в клетках памяти все направления синтеза иммуноглобулинов, за исключением одного, репрессированы. Повторное внедрение антигена для этих клеток служит не сигналом для осуществления выбора синтеза определенного
и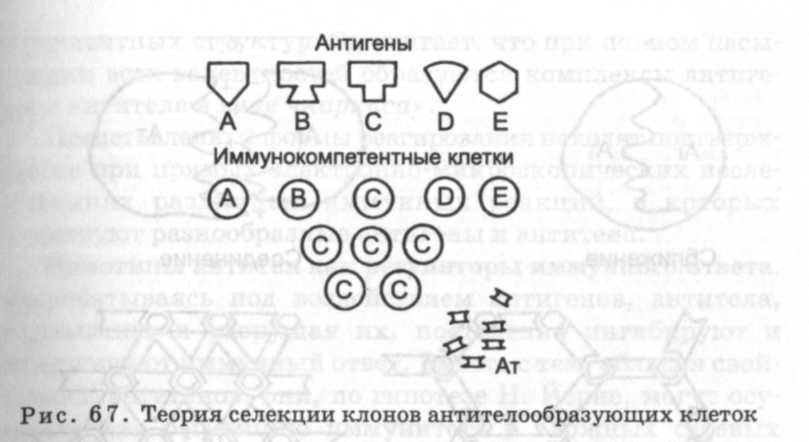 ммуноглобулина,
как для исходных клеток, а директивным
включателем уже детерминированной
дифференциации и пролиферации.
ммуноглобулина,
как для исходных клеток, а директивным
включателем уже детерминированной
дифференциации и пролиферации.
Данная теория охватывает многие стороны иммунитета. Объясняя процесс антителообразования селекцией антигеном соответствующих клонов клеток-предшественников или отбором только некоторого количества из них, она допускает мутационное формирование иммунокомпетентных клеток в онтогенезе. С позиций этой теории стало возможным объяснить состояние толерантности организма к собственным антигенам, под воздействием которых в эмбриональном периоде происходит гибель клеток со специфическими для них рецепторами.
Теория Ф. Бернета стала ключевой и определяющей в развитии современной иммунологии, в частности иммуногенетики.
Теория дерепрессии генов. Теория допускает, что иммунокомпетентная клетка может вырабатывать антитела самой различной специфичности. По минимальным расчетам, ее код содержит информацию, достаточную для формирования 175 ООО антигенсвязывающих центров антител. Однако способность генов детерминировать продукцию такого количества антител репрессирована. Предполагается, что антиген соединяется с ферментом, который обусловливает синтез репрессора и, снимая его действие, индуцирует антителообразование.
Теория взаимодействия антител с антигенами. По вопросу механизма реагирования антигенов с антителами

имеется несколько гипотез. П. Эрлих предполагал, что иммунологические реакции протекают по типу химических и, в частности, представлял, что взаимодействие токсина с антитоксином протекает, как реакция между сильной кислотой и сильной щелочью. Вскоре, однако, выяснилось, что в основе реакции иммунитета лежит адсорбционный процесс, специфичность которого определяется строением детерминантных групп антигенов и соответствующих им по заряду, структуре и конфигурации анти-генсвязывающих центров антител.
Относительно характера соединения антитела с антигеном существуют две гипотезы. По мнению П. Маррека, реакция взаимодействия антигена с антителом обусловлена разноименностью их электрических зарядов (рис. 68). Прочность соединения зависит от количества и полноты совпадения
реагирующих групп антигена и антитела. При оптимальном соотношении антител и антигенов их молекулы в специфических комплексах располагаются в чередующемся порядке и образуют своеобразные «решетки». Л. Полинг рассматривает взаимодействие антитела с антигеном как процесс реагирования двухвалентных и поливалентных структур. Он считает, что при полном насыщении всех валентностей образуются комплексы антигена и антитела в виде «каркаса».
Представленные формы реагирования находят подтверждение при прямых электронно-микроскопических исследованиях различных иммунных реакций, в которых участвуют разнообразные антигены и антитела.
Идиотипы антител как регуляторы иммунного ответа. Вырабатываясь под воздействием антигенов, антитела, связываясь и разрушая их, постепенно ингибируют и прекращают иммунный ответ. Вместе с тем, обладая свойствами антигенов, они, по гипотезе Н. Йерне, могут осуществлять регуляцию иммунитета в сложных сетевых взаимодействиях идиотип-антиидиотип между лимфоцитами различных Т- и В-субпопуляций. Для понимания этого механизма напомним, что каждый специфический антигенсвязывающий фрагмент антитела имеет свой характерный набор идиотипов. В общей массе в организме животных их многие миллионы. Так как огромное разнообразие идиотипов антител, по мнению Н. Йерне, обусловлено многообразием природных антигенных детерминант, то распознающие их лимфоциты должны распознавать также идиотипические детерминанты всех антител, в том числе цитофильных, т. е. самих лимфоцитарных рецепторов. И действительно, так оно и есть. Толерантности к идиотипам у иммунной системы не возникло вследствие ничтожно малых (следовых) их количеств.
И ммунизированное
каким-либо внешним антигеном животное
сначала будет вырабатывать против него
специфические антитела, затем -
антитела к идиотипам антител, а далее
- антитела к антиидиотипическим антителам
и т. д. Такой цепной тип реакций, дающих
«сеть» аутоантител, возникает, когда
большая часть выработанных на антигенный
стимул иммуноглобулинов имеет одинаковый
идиотип. При этом наряду с процессом
выработки аутоантител к доминантному
идиотипу активируются соответствующие
клоны Т-клеток. Те и другие способны
ингибировать или усиливать ответ
лимфоцитов, имеющих рецепторы к данному
идиотипу. Через взаимодействия
идиотип-антиидиотип лимфоциты
функционально связаны между собой
в замкнутую идиотипическую сеть.
ммунизированное
каким-либо внешним антигеном животное
сначала будет вырабатывать против него
специфические антитела, затем -
антитела к идиотипам антител, а далее
- антитела к антиидиотипическим антителам
и т. д. Такой цепной тип реакций, дающих
«сеть» аутоантител, возникает, когда
большая часть выработанных на антигенный
стимул иммуноглобулинов имеет одинаковый
идиотип. При этом наряду с процессом
выработки аутоантител к доминантному
идиотипу активируются соответствующие
клоны Т-клеток. Те и другие способны
ингибировать или усиливать ответ
лимфоцитов, имеющих рецепторы к данному
идиотипу. Через взаимодействия
идиотип-антиидиотип лимфоциты
функционально связаны между собой
в замкнутую идиотипическую сеть.