

2 курс / Гистология / Быков общий
.pdfГлава 3
ЦИТОЛОГИЯ: ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ КЛЕТКИ
ОБЩИЕ ПРИНЦИПЫ СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ КЛЕТКИ И ЕЕ КОМПОНЕНТЫ
Клетка - элементарная структурная, функциональная и генетическая единица в составе всех растительных и животных организмов. Организм взрослого человека состоит примерно из 1013 клеток, которые подразделяют более чем на 200 типов, существенно различающихся своими структурными и функциональными особенностями. Вместе с тем, клетки всех типов характеризуются сходством общей организации и строения важнейших компонентов.
Компоненты клетки. Каждая клетка состоит из двух основных компонентов - ядра и цитоплазмы. В ядре находятся хромосомы, содержащие генетическую информацию, которая в результате процесса транскрипции постоянно избирательно считывается и направляется в цитоплазму, где она контролирует ход многообразных процессов жизнедеятельности клетки, в частности, сбалансированные процессы синтеза, анаболизма (от греч. anabole - повышение), и разрушения, катаболизма (от греч. kataballo - разрушаю). Указанные процессы осуществляются в цитоплазме благодаря взаимодействию ее компонентов.
Компоненты цитоплазмы. Цитоплазма отделена от внешней (для данной клетки) среды внешней клеточной мембраной (плазмолеммой) и содержит органеллы и включения (рис. 3-1), погруженные в гиалоплазму (клеточный матрикс).
Органеллы - постоянно присутствующие в цитоплазме структуры, специализированные на выполнении определенных функций в клетке. Они подразделяются на органеллы общего значения и специальные органеллы.
(1) органеллы общего значения имеются во всех клетках и необходимы для обеспечения их жизнедеятельности. К ним относятся митохондрии, рибосомы, эндоплазмагическая сеть (ЭПС), комплекс Гольджи, лизосомы, порокеисомы, клеточный центр, компоненты цитоскелета;
- 31 -
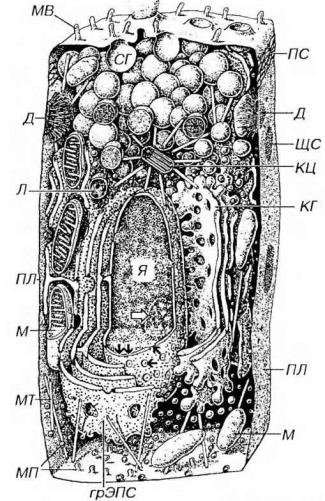
Рис. 3-1. Схема строения клетки (по R.V.Krstic, 1976). Я - ядро; ядрышко показано светлой стрелкой, кариолемма - двойными черными стрелками, ядерные поры - отдельными черными стрелками, М - митохондрии, КГ - комплекс Гольджи, СГ - секреторные гранулы. Л - лизосома, КЦ - клеточный центр, МТ - микротрубочки, ПЛ - плазмолемма, МП - микропиноцитозные пузырьки, МВ - микроворсинки, ПС - плотное соединение, ЩС - щелевое соединение, Д - десмосома.
(2) специальные органеллы имеются лишь в некоторых клетках и обеспечивают выполнение их специализированных функций. К ним относят реснички, жгутики, микроворсинки, миофибриллы, акросому (спермиев). Специальные органеллы образуются в ходе развития клетки как производные органелл общего значения.
- 32 -
В состав многих органелл входит элементарная биологическая мембрана, поэтому органеллы подразделяют также на мембранные и не-мембранные. К мембранным органеллам относятся митохондрии, ЭПС, комплекс Гольджи, лизосомы, пероксисомы, к немембранным - рибосомы, клеточный центр, реснички, микроворсинки, жгутики, компоненты цитоскелета.
Функциональные системы (аппараты) клетки - комплексы органелл,
которые под контролем ядра обеспечивают выполнение важнейших функций клетки. Выделяют: (1) синтетический аппарат;
(1) энергетический аппарат; (3) аппарат внутриклеточного переваривания (эндосомально-лизосомальный); (4) цитоскелет.
Включения - временные компоненты цитоплазмы, образованные в результате накопления продуктов метаболизма клеток. Подразделяются на несколько типов (см. ниже).
Помимо структур цитоплазмы, которые можно четко отнести к органеллам или включениям, в ней имеется огромное количество разно¬образных транспортных пузырьков, обеспечивающих не только перенос веществ между различными компонентами клетки, но и их частичное преобразование (процессинг) благодаря наличию ферментов в мембране, которая образует их стенку.
Мембранные структуры (компоненты) клетки - совокупное на-звание различных структур цитоплазмы и ядра: плазмолеммы, ряда органелл, включений, транспортных пузырьков, а также ядерной оболочки (кариолеммы), в состав которых входят клеточные мембраны. Последние в различных мембранных структурах клетки организованы сходным образом, однако существенно различаются, в первую очередь, составом мембранных белков, определяющим специфику их функций.
Гиалоплазма (клеточный сок, цитозоль, клеточный матрикс) - внутренняя среда клетки, на которую приходится до 55% ее общего объема. Она представляет собой сложную прозрачную коллоидную систему, в которой взвешены органеллы и включения, и содержит различные биополимеры: белки, полисахариды, нуклеиновые кислоты, а также ионы. Претерпевает превращения по типу гельзоль. В гиалоплазме происходит большая часть реакций межуточного обмена.
- 33 -
ПЛАЗМОЛЕММА
Плазмолемма (внешняя клеточная мембрана, цитолемма, плазматическая мембрана) занимает в клетке пограничное положение и играет роль полупроницаемого селективного барьера, который, с одной стороны, отделяет цитоплазму от окружающей клетку среды, а с другой - обеспечивает ее связь с этой средой.
Функции плазмолеммы определяются ее положением и включают:
1)Распознавание данной клеткой других клеток и прикрепление к ним;
2)Распознавание клеткой межклеточного вещества и прикрепление к его элементам (волокнам, базальной мембране);
3)Транспорт веществ и частиц в цитоплазму и из нее (посредством ряда механизмов);
4)Взаимодействие с сигнальными молекулами (гормонами, меди-аторами, цитокинами и др.) благодаря наличию на ее поверхности специфических рецепторов к ним;
5)Движение клетки (образование псевдо-, фило- и ламеллоподий)
- благодаря связи плазмолеммы с сократимыми элементами цитоскелета.
Структура плазмолеммы. Плазмолемма - самая толстая из клеточных мембран (7.5-11 нм); под электронным микроскопом она, как и другие клеточные мембраны, имеет вид трехслойной структуры, представленной двумя электронно-плотными слоями, которые разделены светлым слоем. Ее молекулярное строение описывается жидкостно-мозаичной моделью, согласно которой она состоит из липидного (фосфо-липидного) бислоя, в который погружены и с которым связаны молекулы белков.
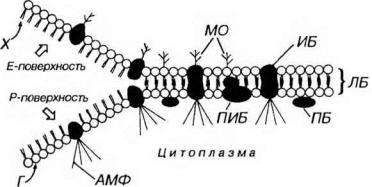
Рис. 3-2. Плазмолемма. ЛБ - липидный бислой: Г - головки (липидных молекул), X - хвосты, ИБ - интегральные белки, ПИБ - полуинтегральные белки, ПБ - периферические белки, МО - молекулы олигосахаридов, связанные с белками и липидами, АМФ - актиновые микрофиламенты, связанные с белками плазмолеммы. Слева показаны поверхности мембраны, выявляемые в результате ее расщепления при замораживании-скалывании.
- 34 -
Липидный бислой представлен преимущественно молекулами фосфатидалхолина (лецитина) и фосфатидилэтаноламина (цефалина), состоящими из гидрофильной (полярной) головки и гидрофобного (неполярного) хвоста. В состав большинства мембран входит также холестерин (холестерол). В мембране гидрофобные цепи обращены внутрь бислоя, а гидрофильные головки - кнаружи (рис.3-2). Состав липидов каждой из половин бислоя неидентичен. Липиды обеспечивают основные физико-химические свойства мембран, в частности, их текучесть при температуре тела. Некоторые липиды (гликолипиды) связаны с олигосахаридными цепями, которые выступают за пределы наружной поверхности плазмолеммы, придавая ей асимметричность. Электронно-плотные слои соответствуют расположению гидрофильных участков липидных молекул.
Мембранные белки составляют более 50% массы мембраны и удерживаются в липидном бислое за счет гидрофобных взаимодействий с молекулами липидов. Они обеспечивают специфические свойства мембраны (типы белков и их содержание в мембране отражают ее функцию) и играют различную биологическую роль (переносчиков, ферментов, рецепторов и структурных молекул). По своему расположению относительно липидного бислоя мембранные белки разделяются на две основные группы - интегральные и периферические (см. рис. 3-2).
Периферические белки непрочно связаны с поверхностью мембраны и обычно находятся вне липидного бислоя.
Интегральные белки либо полностью (собственно интегральные белки), либо частично (полуинтегральные белки) погружены в липидный бислой; часть белков целиком пронизывает всю мембрану (трансмембранные белки). Интегральные белки плазмолеммы хорошо выявляются при использовании метода замораживания-скалывания. При этом плоскость скола обычно проходит через гидрофобную середину бислоя, разделяя его на два листка - наружный и внутренний (см. рис. 3-2). Интегральные белки имеют вид округлых внутримембранных частиц, большая часть которых связана с P-поверхностью (от англ. protoplasmic) - протоплазматической, т.е. ближайшей к цитоплазме поверхности скола (наружной поверхности внутреннего листка), меньшая - на Е-поверхности (от англ. external) - наружной, более близкой к внешней среде поверхности скола (внутренней поверхности наружного листка).
Часть белковых частиц связана с молекулами олигосахаридов (гликопротеины), которые выступают за пределы наружной поверхности
- 35 -
плазмолеммы, другая имеет липидные боковые цепи (липопротеины). Молекулы олигосахаридов связаны также с липидами с составе гликолипидов. Углеводные участки гликолииидов и гликонротеинов придают
поверхности клетки отрицательный заряд и образуют основу так называемого гликокаликса (от греч. glykos - сладкий и calyx - оболочка), который выявляется под электронным микроскопом в виде рыхлого слоя умеренной электронной плотности, покрывающего наружную поверхность плазмолеммы. Эти углеводные участки играют роль рецепторов, обеспечивающих распознавание клеткой соседних клеток и межклеточного вещества, а также адгезивные взаимодействия с ними. В состав гликокаликса некоторые авторы включают, помимо углеводных компонентов, периферические мембранные белки и полуин тегральные белки, функциональные участки которых находятся в над мембранной зоне (например, иммуноглобулины). В гликокаликсе находятся рецепторы гистосовместимости, некоторые ферменты (часть которых может производиться не самой клеткой, а адсорбироваться на ее поверхности), рецепторы гормонов.
Белковые молекулы мозаично распределены в липидном бислое, однако они не жестко фиксированы в нем, а напротив, могут перемещаться в его плоскости. В некоторых условиях определенные белки способны накапливаться в отдельных участках мембраны, образуя агрегаты. Перемещение белковых частиц, но-видимому, не является произвольным, а контролируется внутриклеточными механизмами, в которых участвуют микрофиламенты (см. цитоскелет), прикрепленные к некоторым интегральным белкам, связанным с Р- поверхностъю (см. рис. 3-2).
Мембранный транспорт веществ может включать однонаправленный перенос молекулы какого-то вещества или совместный транспорт двух различных молекул в одном или противоположных направлениях.
Пассивный транспорт включает простую и облегченную дифузию - процессы, которые не требуют затраты энергии. Механизмом простой диффузии осуществляется перенос мелких молекул (например, O2, Н20, С02); этот процесс малоспецифичен и протекает со скоростью, пропорциональной градиенту концентрации транспортируемых молекул но обеим сторонам мембраны. Облегченная диффузия осуществляется через каналы и (или) белкипереносчики, которые обладают специфичностью в отношении транспортируемых молекул. В качестве ионных каналов выступают трансмембранные белки, образующие мелкие водные норы, через которые по электрохимическому градиенту транспортируются мелкие водорастворимые молекулы и ионы. Белки-переносчики также являются трансмембранными белками, которые претерпевают обратимые изменения конформации,
- 36 -
обеспечивающие транспорт специфических молекул через плазмолемму. Они функционируют в механизмах как пассивного, так и активного транспорта.
Активный транспорт является энергоемким процессом, благодаря которому перенос молекул осуществляется с помощью белков-переносчиков против электрохимического градиента. Примером механизма, обеспечивающего противоположно направленный активный транспорт ионов, служит натриевокалиевый насос (представленный белком-пере- носчиком Na+-К+-АТФазой), благодаря которому ионы Na+ выводятся из цитоплазмы, а ионы К+ одновременно переносятся в нее. Этот механизм обеспечивает поддержание постоянства объема клетки (путем регуляции осмотического давления), а также мембранного потенциала. Активный транспорт глюкозы в клетку осуществляется белком-переносчиком и сочетается с однонаправленным переносом иона Na+.
Облегченный транспорт ионов опосредуется особыми трансмембранными белками - ионными каналами, обеспечивающими избирательный перенос определенных ионов. Эти каналы состоят из собственно транспортной системы и воротного механизма, который открывает канал на некоторое время в ответ на (а) изменение мембранного потенциала, (б) механическое воздействие (например, в волосковых клетках внутреннего уха), (в) связывание лиганда (сигнальной молекулы или иона).
Эндоцитоз. Транспорт макромолекул в клетку осуществляется с помощью механизма эндоцитоза (от греч. endo - внутрь и cytos - клетка). Материал, находящийся во внеклеточном пространстве, захватывается в области впячивания (инвагинации) плазмолеммы, края которого смыкаются с формированием эндоцитозного пузырька или эндосомы - мелкого сферического образования, герметически окруженного мембраной (рис. 3-3 и 3-5). Далее содержимое эндосомы подвергается внутриклеточной переработке (процессингу). В частности, в эндосоме в условиях закисления среды происходит отделение лиганда от рецептора (последний в дальнейшем используется повторно) - см. ниже. Разновидностями эндоцитоза служат пиноцитоз и фагоцитоз.
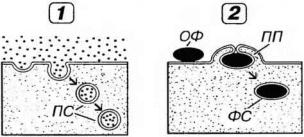
Рис. 3-3. Пиноцитоз (1) и фагоцитоз (2). ПС - пиносомы, ОФ - объект фагоцитоза, ПП - псевдоподии, ФС - фагосома.
- 37 -
Пиноцитоз (от греч. pinein - нить и cytos - клетка) - захват и поглощение клеткой жидкости и (или) растворимых веществ; подразделяется на макропиноцитоз (диаметр эндосом 0.2-0.3 мкм) и микропиноцитоз (диаметр эндосом - 70-100 нм).
Фагоцитоз (от греч. phagein - поедать и cytos - клетка) - захват и поглощение клеткой плотных, обычно крупных (размером более 1 мкм) частиц; обычно сопровождается образованием выпячиваний цитоплазмы - псевдоподий, охватывающих объект фагоцитоза и смыкающихся над ним (см. рис. 3-3).
Рецепторно-опосредованный эндоцитоз. Эффективность эндоцитоза существенно увеличивается, если он опосредован мембранными рецепторами, которые связываются с молекулами поглощаемого вещества или молекулами, находящимися на поверхности фагоцитируемого объекта - лигандами (от лат. ligare - связывать). В дальнейшем (после поглощения вещества) комплекс рецептор-лиганд расщепляется, и рецепторы могут вновь возвратиться в плазмолемму.
Примером рецепторно-опосредованного взаимодействия может служить фагоцитоз лейкоцитом бактерии (см. рис. 7-8). Поскольку на плазмолемме лейкоцита имеются рецепторы к иммуноглобулинам (антителам), скорость фагоцитоза резко возрастает, если поверхность бактерии покрыта антителами (опсонинами - от греч. opson - приправа).
Окаймленные пузырьки и ямки. Рецепторы макромолекул в плазмолемме, перемещаясь латерально по клеточной поверхности, могут, связывая свои лиганды, накапливаться в области формирующихся эндоцитозных ямок. Очень часто вокруг таких ямок и образующихся из них пузырьков со стороны цитоплазмы собирается сетевидная оболочка из белка клатрина, которая на срезах имеет вид щетинистой каемки (рис. 3-4). В покрытых клатриновой оболочкой (окаймленных) ямках рецепторные белки мембраны вытесняют все остальные; таким образом ямки действуют как приспособления для накопления и сортировки молекул. Этим механизмом достигается и значительная экономия в ходе процесса эндоцитоза: для поглощения определенного количества молекул лиганда требуется значительно меньше пузырьков, чем было бы в случае диффузного распределения комплексов рецептор-лиганд.
Окаймленная ямка достигает своего максимального размера (около 0. 3 мкм) в течение 1 мин и превращается в окаймленный пузырек. Его содержимое может подвергаться процессингу лишь после того, как через несколько секунд он утратит клатриновую оболочку. Если она сохраняется, пузырек не способен сливаться с другими структурами (ана-
- 38 -
логичными пузырьками, лизосомами), и его содержимое остается неизмененным. Окаймленные эндоцитозные пузырьки транспортируют иммуноглобулины, белки желточных включений (в цитоплазму овоцитов), факторы роста, липопротеины низкой плотности (ЛНП). Некоторые транспортные мембранные пузырьки в цитоплазме окружены неклатриновой белковой оболочкой.
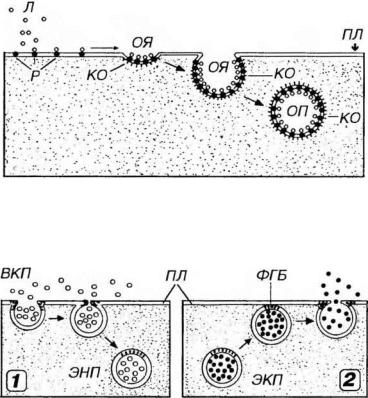
Рис. 3-4. Рецепторно-опосредованный эндоцитоз. ПЛ - плазмолемма, Л - лиганд. Р - рецепторы, ОЯ - окаймленная ямка, ОП - окаймленный пузырек, КО - клатриновая оболочка.
Рис. 3-5. Эндоцитоз (1) и экзоцитоз (2). ВКП - внеклеточное пространство, ПЛ - плазмолемма, ЭНП - эндоцитозный пузырек, ЭКП - экзоцитоэный пузырек, ФГБ - фузогенные белки.
Нарушение транспорта ЛНП описанным механизмом при врожденном наследственном заболевании - семейной гиперхолестеринемии - обусловлено отсутствием или наличием дефектных рецепторов ЛНП, неспособных связывать лиганд или накапливаться в окаймленных ямках. При этом поглощение клетками холестерина, поступающего с ЛНП, ослаблено, а его уровни в крови резко повышены, вызывая быстрое развитие атеросклероза и смерть больных в молодом возрасте от ишемической болезни сердца.
- 39 -
Экзоцитоз (от греч. ехо - наружу и cytos - клетка) - процесс, обратный эндоцитозу, при котором мембранные экзоцитозные пузырьки приближаются к плазмолемме и сливаются с ней своей мембраной, которая встраивается в плазмолемму. При этом содержимое пузырьков (продукты собственного синтеза клетки или транспортируемые ею молекулы, непереваренные и вредные вещества и др.) выделяется во внеклеточное пространство (см. рис. 3-5).
Судьба выделяемых экзоцитозом синтезированных клеткой молекул неодинакова: (1) прикрепляясь к клеточной поверхности, они могут становиться периферическими белками (например, антигенами); (2) они могут войти в состав межклеточного вещества (например, коллаген и гликозаминогликаны; (3) попадая во внеклеточную жидкость, они могут выполнять роль сигнальных молекул (гормоны, цитокины).
Трансцитоз (от лат. trans - сквозь, через и греч. cytos - клетка) процесс, характерный для некоторых типов клеток, объединяющий признаки эндоцитоза и экзоцитоза. На одной поверхности клетки формируется эндоцитозный пузырек, который переносится к противоположной поверхности клетки и, становясь экзоцитозным пузырьком, выделяет свое содержимое во внеклеточное пространство. Процессы трансцитоза протекают очень активно в цитоплазме плоских клеток, выстилающих сосуды (эндотелиоцитах), особенно в капиллярах. В этих клетках пузырьки, сливаясь, могут образовывать временные трансцеллюлярные каналы, через которые транспортируются водорастворимые молекулы.
Ход образования эндоцитозных пузырьков опосредуется особыми (фузогенными - от лат. fusio - слияние) мембранными белками, которые концентрируются в участках инваганации плазмолеммы. Эти же белки при экзоцитозе способствуют слиянию мембраны пузырька с плазмолеммой (см. рис. 3-5). Важную роль в процессах эндоцитоза и экзоцитоза играют элементы цитоскелета, в частности, микрофиламенты и микротрубочки (см. ниже).
Баланс процессов эндоцитоза и экзоцитоза. Эндоцитоз вследствие постоянной отпшуровки пузырьков с поверхности плазмолеммы должен приводить к уменьшению ее площади при одновременном увеличении объема клетки. Так, например, в макрофагах за 1 ч за счет эндоцитоза вносится до 25% объема цитоплазмы, а за 0.5 ч общая площадь поверхности эндоцитозных пузырьков составляет 100% площади плазмолеммы. При экзоцитозе, напротив, постоянно происходит увеличение площади плазмолеммы вследствие встраивания в нее мембраны экзоцитозных пузырьков. Так, в секреторной клетке ацинуса поджелудочной железы совокупная площадь мембраны секреторных гранул в 30 раз больше, чем поверхность плазмолеммы.
- 40 -