

2 курс / Гистология / Быков общий
.pdfПроцесс ороговения эпителия заключается в превращении его живых эпителиальных клеток в роговые чешуйки - механически прочные и химически устойчивые постклеточные структуры, образующие в совокупности роговой слой эпителия, который обладает защитными свойствами. Хотя собственно формирование роговых чешуек происходит в наружных отделах зернистого слоя или в блестящем слое, синтез веществ, обеспечивающих ороговение, осуществляется еще в шиповатом слое.
Основные процессы, происходящие в клетках в ходе ороговения:
(1)изменение формы - клетки резко уплощаются, приобретая шестиугольную форму;
(2)сборка и стабилизация пучков кератиновых промежуточных филаментов, образующих сети в цитоплазме и составляющих 80% ее массы. Процесс организации сетей филаментов с формированием их крупных пучков (макрофиламентов) обеспечивается филаггрином, который по его завершении разрушается. Последующая стабилизация системы филаментов осуществляется путем формирования межмолекулярных дисульфидных связей. Кератиновые филаменты присутствуют в клетках, начиная с базального слоя, филаггрин в виде неактивного предшественника (профилаприна) впервые обнаруживается в кератогиалиновых гранулах зернистого слоя;
(3)образование оболочки роговой чешуйки - плотной, механически прочной и химически устойчивой белковой структуры - происходит путем упорядоченного отложения ряда белков (инволюкрина, цистатина-α, лорикрина
идр.) на внутренней поверхности плазмолеммы с образованием между ними многочисленных ковалентных связей. Сборка оболочки роговой чешуйки катализируется ферментом трансглутаминазой (который, как и инволюкрин - его главный субстрат и основной компонент оболочки - впервые обнаруживается в шиповатом слое).
(4)полное ферментное разрушение остальных структур цитоплазмы и
ядра;
(5)дегидратация цитоплазмы (с потерей 70% массы клетки).
5) Роговой слой - наиболее поверхностный - имеет максимальную толщину в эпителии кожи (эпидермисе) в области ладоней и подошв. Он образован плоскими роговыми чешуйками с резко утолщенной плазмолеммой (оболочкой), не содержащими ядра и органелл и заполненными сетью из толстых пучков кератиновых филаментов, погруженных в плотный матрикс. Роговые чешуйки в течение определенного времени сохраняют связи друг с другом и удерживаются в составе пластов благодаря частично сохраненным десмосомам, а также взаимному проникно-
- 141 -

вению бороздок и гребешков, образующих ряды на поверхности соседних чешуек. В наружных частях слоя десмосомы полностью разрушаются, и роговые чешуйки слущиваются с поверхности эпителия.
Многослойный плоский неороговевающий эпителий покрывает поверхность роговицы глаза, конъюнктивы, слизистых оболочек полости рта (частично), глотки, пищевода, влагалища, влагалищной части шейки матки, части мочеиспускательного канала. Он образован тремя слоями клеток: (1) базальным, (2) шиповатым (промежуточным) и (3) поверхностным (рис. 5-12 и
5-13).
Рис. 5-12. МНОГОСЛОЙНЫЙ ПЛОСКИЙ неороговевающий эпителий (влагалище) - вид на гистологическом препарате. БМ - базальная мембрана. БС - базальный слой, ШС - шиповатый слой, ПС - поверхностный слой. Десквамирующая клетка поверхностного слоя показана стрелкой.
1)Базальный слой аналогичен по строению и функции соответствующему слою ороговевающего эпителия.
2)Шиповатый (промежуточный) слой образован крупными полигональными клетками, которые по мере приближения к поверхностному слою уплощаются. Их цитоплазма заполняется многочисленными тонофиламентами, которые располагаются в ней диффузно, не образуя крупных пучков. В клетках наружных отделов этого слоя накапливается кератогиалин в виде мелких округлых гранул.
-142 -
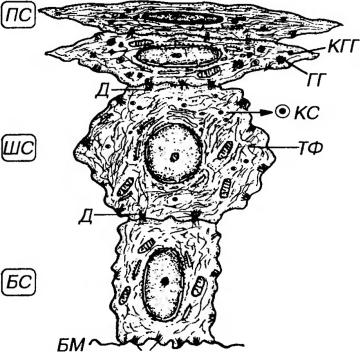
3) Поверхностный слой нерезко отделен от шиповатого. Он образован уплощенными клетками, содержащими рыхло распределенные цитокератиновые филаменты, которые по химическому составу отличаются от таковых в роговых чешуйках. Содержание органелл снижено по сравнению с таковым в клетках шиповатого слоя, плазмолемма утолщена, межклеточные пространства редуцированы. Ядро - светлое (везикулярное) или темное, с плохо различимыми гранулами хроматина (пикнотическое). Механизмом десквамации клетки этого слоя постоянно удаляются с поверхностности эпителия.
Рис. 5-13. Ультраструктурная организация многослойного плоского неороговевающего эпителия. БМ - базальная мембрана, БС - базальный слой, ШС - шиповатый слой, ПС - поверхностный слой, ТФ - тонофиламенты, КС - кератиносомы, КГГ - кератогиалиновые гранулы, ПД - полудесмосомы, Д - десмосомы, ГГ - гранулы гликогена.
Многослойный кубический эпителий в организме человека встречается редко. Он сходен по строению с многослойным плоским эпителием, но клетки поверхностного слоя в нем имеют кубическую форму. Такой эпителий образует стенку крупных фолликулов яичника, выстилает протоки потовых и сальных желез кожи.
Многослойный призматический эпителий, как и многослойный кубический, у человека встречается редко. Он выстилает некоторые
- 143 -

участки мочеиспускательного канала, крупные выводные протоки слюнных и молочных желез (частично). Такой эпителий обнаруживается в участках резкого перехода многослойного плоского эпителия в однослойный многорядный и образует узкую зону между этими эпителиями (например, в глотке и гортани).
Переходный эпителий - особый вид многослойного эпителия, который выстилает большую часть мочевыводящих путей (чашечки, лоханки, мочеточники и мочевой пузырь, часть мочеиспускательного канала), отчего его называют также уротелием. Форма клеток этого эпителия и толщина всего эпителиального пласта зависят от функционального состояния (степени растяжения) органа. Переходный эпителий образован тремя слоями клеток: базальным, промежуточным и поверхностным (рис. 5-14).
Рис. 5-14. Переходный эпителий. 1 - в состоянии покоя, 2 - в состоянии растяжения, 3 - участок цитоплазмы апикальной части фасеточной клетки на срезе, 4 - то же в объемном изображении, 5 - внутри мембранная частица (ВМЧ), входящая в состав пластинок плазмолеммы (ПП) фасеточной клетки. БМ - базальная мембрана, БС - базальный слой, ПРС - промежуточный слой, ПОС - поверхностный слой (образован фасеточными клетками), ДП - дисковидные пузырьки, МФ - микрофиламенты.
(1) базальный слой образован мелкими клетками, имеющими на срезе преимущественно треугольную форму и своим широким основанием прилежащими к тонкой базальной мембране;
- 144 -
(2)промежуточный слой состоит из удлиненных клеток, более узкой частью направленных к базальному слою и черепицеобразно накладывающихся друг на друга;
(3)поверхностный слой образован крупными одноядерными полиплоидными или двуядерными поверхностными (фасеточными) клетками, которые в наибольшей степени изменяют свою форму при растяжении эпителия (от округлой до плоской). Этому способствует формирование в апикальной части цитоплазмы этих клеток в состоянии покоя многочисленных инвагинаций плазмолеммы и особых дисковидных (веретеновидных на срезах) пузырьков (длиной 0.3-0.8 мкм и шириной 0.12-0.18 мкм) - резервов плазмолеммы, которые встраиваются в нее по мере растяжения клетки. Формированию инвагинаций плазмолеммы способствуют многочисленные микрофиламенты, которые прикрепляются к ее особым участкам - пластинкам плазмолеммы.
Пластинки плазмолеммы - утолщенные, сравнительно ригидные и малопроницаемые для воды полигональные участки апикальной плазмолеммы площадью 0.05-0.25 мкм2, которые в совокупности занимают до 75% ее поверхности. Пластинки содержат скопления внутримембранных белковых частиц размером около 12 нм, каждая из которых образована шестью субъединицами диаметром 5 нм. Пластинки плазмолеммы располагаются в виде "булыжной мостовой" (за тем лишь исключением, что каждая из них представляет собой не выпячивание, а вдавление на ее поверхности). Они разделены более гибкими участками плазмолеммы, не содержащими белковых частиц, которые способствуют образованию складок плазмолеммы. Мембрана, образующая пластинки плазмолеммы, собирается в комплексе Гольджи и транспортируется в апикальную плазмолемму посредством дисковидных пузырьков. Предполагают, что наличие указанных пластинок в апикальной плазмолемме поверхностных клеток и плотных соединений между латеральными поверхностями этих клеток обеспечивает непроницаемость переходного эпителия для воды. Эго свойство данного эпителия имеет важнейшее функциональное значение, поскольку благодаря ему гипертоническая моча (накапливающаяся в мочевом пузыре) не разводится изотонической жидкостью из кровеносных сосудов подлежащей соединительной ткани собственной пластинки слизистой оболочки.
Альтернативные представления о строении переходного эпителия основаны на данных некоторых исследователей, согласно которым клетки его промежуточного и даже поверхностного слоев своими тонкими отростками контактируют с базальной мембраной. В соответствии с такими представлениями, переходный эпителий следует считать не многослойным, а особым видом однойслойного многорядного.
- 145 -
ГИСТОГЕНЕТИЧЕСКАЯ КЛАССИФИКАЦИЯ ЭПИТЕЛИЕВ
Гисто генетическая классификация эпителиев разработана акад.
Н.Г.Хлопиным и выделяет пять основных типов эпителия, развивающихся в эмбриогенезе из различных тканевых зачатков (см. ниже).
Гистогенетическая классификация эпителиев (по Н.Г.Хлопину)
Гистогенетический тип эпителия |
Эмбриональные зачатки - |
|
|
|
источники развития эпителия |
1. |
Эпидермальный |
эктодерма, прехордальная пластинка |
2. |
Энтеродермальный |
кишечная энтодерма |
3. |
Целонефродермальный |
целомическая выстилка, нефротом |
4. |
Ангиодермальный |
ангиобласт |
5. |
Эпендимоглиальный |
нервная трубка |
1. Эпителии эпидермального типа развиваются из эктодермы и прехордальной пластинки, которая также обладает эктодермальной детерминацией. Самым типичным представителем данного тканевого ти-а служит эпителий кожи (эпидермис), наиболее общей функцией - защитная, самым характерным морфологическим признаком - многослойность или многорядность (последняя при повреждении эпителия может сменяться многослойностью).
К эпидермальному типу помимо эпидермиса относят эпителии, вытилающие полость рта, глотки, пищевода, воздухоносных путей и респираторного отдела легких, мочевыводяншх путей, влагалища, роговицу глаза. Эпителии эпидермального типа образуют железы кожи и железы, связанные с указанными выше слизистыми оболочками, функционально ведущую ткань крупных слюнных и молочных желез, ряда эндокринных желез (аденогипофиза, щитовидной и околощитовидных желез), эпите-
- 146 -
лиальную основу тимуса. Некоторые авторы относят эпителии - производные прехордальной пластинки не к эпидермальному, а к энтеродермальному тканевому типу.
2.Эпителии энтеродермального типа (от греч. enteron - кишка)
являются производными кишечной энтодермы. Самым типичным представителем этих эпителиев служит эпителий кишки, наиболее типичными функциями - всасывание и (или) секреция, самым характерным морфологическим признаком - однослойность. Эпителии энтеродермального типа образуют выстилку желудочно-кишечного тракта, а также все связанные с ней железы (от самых мелких, расположенных в его стенке, до самых крупных - печени и поджелудочной железы).
3.Эпителии целонефродермалъного типа (от греч. coelom - целом,
полость тела и nephros - почка) развиваются из целомической выстилки и нефротома. Их строение весьма разнообразно; наиболее часто они образуют однослойные однорядные выстилки, но встречаются также однослойные многорядные (например, в семявыносящих путях, предстательной железе) и многослойные (например, в фолликулах яичника). Функции этих эпителиев связаны с процессами секреции, экскреции и всасывания; они выполняют также барьерную функцию. К эпителиям целонефродермалъного типа относят мезотелий (однослойный плоский эпителий, образующий выстилку полостей тела и входящий в состав серозных оболочек), эпителий нефрона, семявыносящих путей, выстилки маточных труб, матки, шеечного канала. Эти эпителии входят в состав яичника (фолликулярные клетки, зернистые лютеоциты) и яичка (сустентоциты, или клетки Сертоли), образуют функционально ведущую ткань предстательной железы и коркового вещества надпочечника.
4.Эпителий ангиодермалъного типа (от греч. angion - сосуд) являются производными особого эмбрионального зачатка ангиобласта, который располагается среди клеток мезенхимы и дает начало выстилке кровеносных и лимфатических сосудов, а также сердца. Единственным представителем данного типа служит эндотелий, который обычно является однослойным плоским эпителием. Исключение составляют выстилки сосудов с особыми функциональными свойствами - (а) посткапиллярных вену л с высоким (кубическим) эндотелием в органах иммунной системы, (б) синусов селезенки с палочковидными эндотелиальными клетками. Функции эндотелия многообразны: он участвует в процессах двустороннего транспорта веществ и клеток между кровью и другими тканями, регуляции свертывания крови и
-147 -
тонуса сосудов, секретирует биологически активные вещества, обеспечивает новообразование сосудов (ангиогенез).
5. Эпителии эпендимоглиального типа - особые ткани нейрального происхождения, выполняющие в нервной системе опорную, разграничительную и секреторную функции, имеющие строение эпителия и относящиеся к так называемой эпендимной глии, давшей им свое название (от греч. ependyma - верхняя одежда и glia, или neuroglia - нервный клей, т.е. ткань, связывающая элементы нервной системы). Этот эпителий выстилает центральный канал спинного мозга и желудочки головного мозга, участвуют в выработке спинномозговой жидкости (см. главу 14). Он образован одним слоем кубических или призматических клеток, которые часто имеют реснички на апикальной поверхности и длинный базальным отросток. По мнению некоторых авторов, эпителии эпендимоглиального типа образуют также выстилки мозговых оболочек, входят в состав органов чувств (равновесия, слуха, зрения) и, возможно, нервных стволов (формируя периневрий).
ЖЕЛЕЗЫ
Железы выполняют секреторную функцию, вырабатывая и выделяя разнообразные продукты (секреты), обеспечивающие различные функции организма. Большинство желез образовано эпителиальной тканью (железистым эпителием), хотя той или иной способностью к секреции обладают все ткани.
Строение и гистофизиология желез
Железистые клетки (гландулоциты - от лат. glandula - железа и kytos, или cytos - клетка) специализированы на выработке секретов, поэтому для них характерны все признаки клеток с активно протекающими синтетическими процессами (рис. 5-15).
Ядро гландулоцитов - обычно крупное, с преобладанием эухроматина, одним или несколькими крупными ядрышками. Его положение в клетке может изменяться в разные фазы секреторного цикла (см. ниже), например, оно может смещаться к базальному полюсу при накоплении секреторных гранул в апикальном.
Цитоплазма гландулоцитов содержит мощно развитая синтетический аппарат, морфологические и функциональные особенности которого зависят от химической природы продуцируемого секрета (см. главу 3). Процессы синтеза и
- 148 -

выделения веществ требуют значительного количества энергии, которая вырабатывается большим числом
митохондрий, находящихся в цитоплазме. Избыток синтезируемых продуктов часто удаляется внутриклеточным механизмом кринофагии, что обусловлено хорошим развитием лизосомального аппарата. Распределение органелл в цитоплазме клеток желез неравномерно в связи с их выраженной полярностью.
Рис. 5-15. Структурно-функциональная организация железистой клетки в секреторном цикле. 1 - фаза поглощения исходных веществ, 2 - фаза синтеза секрета, 3 - фаза накопления синтезированного продукта, 4 - фаза выведения секрете. МТХ - митохондрии, КГ - комплекс Гольджи, НГнезрелые гранулы, СГ - секреторные гранулы (зрелые).
Секреторный цикл. Процесс секреции в железистых клетках протекает циклически и включает четыре фазы, которые могут в различной степени взаимно перекрываться: (1) фазу поглощения исходных веществ, (2) фазу синтеза секрета, (3) фазу накопления синтезированного продукта, (4) фазу выведения секрета (см. рис. 5-15).
1. Фаза поглощения исходных веществ, служащих субстратами для синтеза секреторного продукта, обеспечивается высокой активностью транспортных механизмов, связанных с плазмолеммой базального полюса клетки, через который указанные вещества поступают из крови. В некоторых клетках субстраты для синтеза метут в значительных количествах запасаться в цитоплазме (например, в виде липидных капель в стероид-продуцирующих клетках).
- 149 -
2.Фаза синтеза секрета связана с процессами транскрипции и трансляции, деятельностью грЭПС и комплекса Гольджи (для белковых секретов), аЭПС и митохондрий с тубулярно-везикулярными крис¬ами (для стероидных веществ). Синтезированный продукт в комплексе Гольджи или внутри секреторных гранул нередко претерпевает посттрансляционные изменения, обусловленные действием различных ферментов ("дозревает").
3.Фаза накопления синтезированного продукта в цитоплазме железистых клеток обычно проявляется нарастанием содержания секреторных гранул, которые в некоторых случаях могут укрупняться, сливаясь друг с другом. Переполнению цитоплазмы секреторными гранулами препятствует механизм лизосомального разрушения их избытка - кринофагия. Скопления гранул располагаются преимущественно у апикального полюса клеток экзокринных желез и у базального - в клетках эндокринных желез (см. ниже). Некоторые виды синтезированных продуктов (например, стероидные гормоны) не накапливаются в цитоплазме железистых клеток, а по мере образования, повидимому, сразу же из нее выводятся.
4.Фаза выведения секрета может осуществляться несколькими механизмами (см. ниже). Наиболее часто происходит экзоцитоз содержимого секреторных гранул путем слияния мембраны их гранул с плазмолеммой и выделения синтезированного продукта за пределы клетки. Встроенная в плазмолемму мембрана секреторных гранул затем отделяется из нее в цитоплазму механизмом эндоцитоза и возвращается в комплекс Гольджи для повторного использования (реутилизации, или рециклирования). Некоторые секреты (например, стероидные или тиреоидньге гормоны) выделяются из клетки механизмами диффузии.
Классификация желез
Существует несколько классификаций желез, которые основаны на учете различных признаков. Железы подразделяются:
1)по числу клеток (рис. 5-16) - на одноклеточные (например, бокаловидные клетки, клетки диффузной эндокринной системы) и многоклеточные (большинство желез);
2)по уровню организации - на входящие в состав различных органов в качестве их компонентов (например, железы слизистых оболочек) или
-150 -