

2 курс / Гистология / Быков общий
.pdfсо временем суток, характером и тяжестью выполняемой работа, приемом пищи и другами факторами.
Концентрация лейкоцитов у детей в норме меняется в зависимости от возраста: у новорожденного она равняется 10 000-30 000/мкл (в среднем, 20 000/мкл), на 4-й день снижается до 12 000, к 4-м годам составляет 8000/мкл. Уровня, характерного для взрослого, этот показатель достигает примерно к 1214 годам. С возрастом происходят изменения не только количества, но и качественного состава лейкоцитов.
Лейкоцитоз - увеличение концентрации лейкоцитов в крови - обычно является следствием их усиленного выброса из костного мозга в связи с возросшей потребностью, определяющейся повышенной гибелью (чаще всего при инфекционных и воспалительных заболеваниях).
Лейкопения - снижение концентрации лейкоцитов в крови - как правило, служит результатом подавления их образования в костном мозге (в результате тяжелых инфекционных процессов, токсических состояний, облучения).
Движения лейкоцитов можно разделить на пассивные и активные. Пассивное движение обусловлено переносом лейкоцитов с током крови. Активные движения совершаются благодаря наличию в цитоплазме лейкоцитов многочисленных актиновых микрофиламентов и связанных с ними белков; они осуществляются с затратами энергии.
Миграция лейкоцитов из сосудистого русла в периферические ткани служит важнейшим этапом и условием осуществления функций различными видами этих клеток. Этот процесс происходит в микроциркуляторном русле и наиболее активно протекает, как правило, на уровне посткапиллярных венул. Он включает закономерную серию (каскад) адгезивных взаимодействий между лейкоцитами и клетками эндотелиальной выстилки сосудов. Эти взаимодействия опосредуются последовательной экспрессией на поверхности лейкоцитов и эндотелия характерных комбинаций адгезивных молекул и включают несколько стадий (рис. 7-7):
1.Случайные контактные взаимодействия между лейкоцитами и эндотелиальными клетками - осуществляются постоянно в физиологических условиях с дальнейшим возвращением лейкоцитов в кровоток или переходом к последующим стадиям взаимодействия. Могут усиливаться при изменении условий кровотока, например, при его замедлении;
2.Качение (rolling) лейкоцитов по поверхности эндотелия обусловлено их транзиторной адгезией к выстилке сосуда (посредст-
-181 -
вом адгезивных белков селектинов, экспрессируемых на лейкоцитах и эндотелии). Оно отражает неустойчивое равновесие локальных сил прикрепления (адгезии) лейкоцита к эндотелию и гемодинамических сил, отрывающих его от стенки сосуда. При качении скорость перемещения лейкоцита уменьшается примерно в 100 раз по сравнению с таковой в кровотоке. Начальная активация эндотелия сосуда (расположенного вблизи очага повреждения) цитокинами и медиаторами воспалительных реакций вызывает повышение экспрессии на его поверхности адгезивных молекул и вовлечение в процесс качения все большего числа лейкоцитов. Этот этап является обратимым (быстро блокируется при инактивации селектинов);
3. Остановка качения лейкоцитов, их активация и прочное прикрепление к эндотелию обусловлены продолжающейся стимуляцией эндотелия и лейкоцитов цитокинами, (в том числе недавно открытым классом хемотаксических цитокинов - хемокинов), продуктами повреждения тканей и хемоаттрактантами. Лейкоциты распластываются на поверхности эндотелия и по мере усиления адгезивных взаимодействий прочно прикрепляются к выстилке сосуда. Этот этап, как и последующие, является необратимым и опосредуется адгезивными белками интегринами, селектинами и представителями иммуноглобулиноподобных адгезивных белков.
2. Миграция адгезированных лейкоцитов через эндотелий
осуществляется путем размыкания ими соединений между эндотелиоцитами и проникновения в межклеточные промежутки. При этом лейкоциты плотно прилегают к поверхности клеток эндотелия, последовательно осуществляют с ними ряд адгезивных взаимодействий и всегда целиком заполняют межклеточное пространство, не увеличивая общей проницаемости сосуда. Перемещаясь, лейкоциты сначала образуют псевдоподию, а в дальнейшем их цитоплазма постепенно "перетекает" в сторону сформированного выпячивания. Базальная мембрана не служит препятствием на пути миграции лейкоцитов.
5. Миграция лейкоцитов за пределами сосуда происходит благодаря сократительной активности элементов их цитоскелета и их многочисленным обратимым адгезивным взаимодействиям с клетками различных тканей (в первую очередь, соединительной) и компонентами межклеточного вещества (базальными мембранами, волокнами, гликопротеинами, протеогликанами и др.). Направленность движений лейкоцитов обусловлена их хемотаксисом (перемещением по градиенту привлекающего химического вещества - хемоаттрактанта) и характером адгезивных взаимодействий.
- 182 -
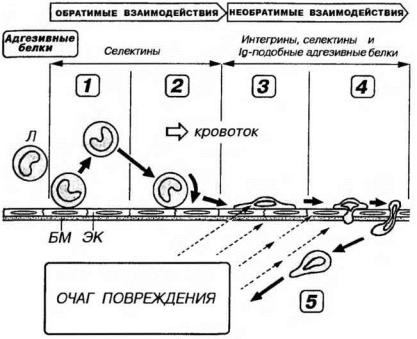
Рис. 7-7. Миграция лейкоцитов из сосудистого русла в ткани. 1 - случайные контактные взаимодействия между лейкоцитами (Л) и эндотелиальными клетками (ЭК) с дальнейшим возвращением Л в кровоток. 2 - качение Л по поверхности ЭК. 3 - остановка качения Л и их активация, прочное прикрепление к ЭК с распластыванием на них. 4 - миграция Л через пласт ЭК и базальную мембрану (БМ). 5 - миграция Л за пределами сосуда. Активация ЭК, Л и направленная миграция последних опосредуются химическими сигналами, исходящими из очага повреждения (прерывистые стрелки).
Способность к целенаправленным движениям обеспечивает перемещение лейкоцитов в окружающую сосуды соединительную ткань, миграцию в органы иммунной системы, проникновение в эпителиальные выстилки и накопление в очагах повреждения тканей и инвазии микробов.
Избирательность миграции лейкоцитов в ткани. В физиологических условиях в отсутствие стимуляции активность миграции лейкоцитов различных видов за пределы сосудистого русла существенно различается. Она сравнительно невелика у нейтрофильных, эозинофильных и базофильных гранулоцитов (см. ниже), резко возрастая при воспалении (см. главу 10). В то же время нестимулированная миграция моноцитов и лимфоцитов происходит очень активно (см. ниже, а так-
- 183 -
же главу 8). Регуляция потока лейкоцитов, мигрирующих за пределы сосудистого русла, имеет некоторые отличия в органах иммунной системы и других тканях и органах, что в значительной мере обусловлено особенностями их сосудов, в частности, посткапиллярных венул.
Посткапиллярные венулы в органах иммунной системы выстланы особым высоким (кубическим) эндотелием, который контролирует перемещение лимфоцитов из кровотока в эти органы. Этот эффект достигается путем экспрессии на поверхности клеток эндотелия специальных адгезивных молекул - адрессинов (различных в отдельных иммунных органах), которые благодаря специфическому взаимодействию с хоминг-рецепторами на лимфоцитах (от англ. homing - возвращение домой) указывают последним направление миграции.
Посткапиллярные венулы в органах, не относящихся к иммунной системе, выстланы обычным (плоским) эндотелием, который обладает способностью избирательно контролировать активность миграции лейкоцитов. Это осуществляется путем экспрессии на его поверхности специфических комбинаций адгезивных молекул (эндотелиального "почтового индекса"), предпочтительно связывающихся с поверхностью лейкоцитов того или иного вида. Указанный процесс зависит от характера стимуляции эндотелия и лейкоцитов цитокинами, хемоаттрактантами и другими веществами, выделяющимися из очага воспаления. Более того, экспрессия адгезивных молекул на эндотелии количественно и качественно меняется во времени, что обусловливает смену потоков лейкоцитов отдельных видов, которые устремляются в участок повреждения тканей на разных сроках после его возникновения.
Нарушения подвижности лейкоцитов (вследствие дефектов цитоскелета),
их способности к адгезивным взаимодействиям или целенаправленному движению (обычно в результате наследственной патологии) обусловливают ряд клинических синдромов, связанных с тяжелыми инфекционными поражениями организма.
Классификация лейкоцитов
Классификация лейкоцитов основана на ряде признаков, из которых ведущим служит присутствие в их цитоплазме специфических гранул. На основании этого признака все лейкоциты разделяют на гранулоциты и агранулоциты.
- 184 -
Гранулоциты (зернистые лейкоциты) характеризуются наличием в их цитоплазме специфических гранул, обладающих различной окраской (базофильной, оксифильной или нейтрофильной). Это, в своюочередь, позволяет подразделять гранулоциты на базофильные, оксифильные
(эозинофильные) и нейтрофильные. В гранулоцитах присутствует и второй тип гранул - неспецифические, или азурофильные (окрашиваются азуром и являются лизосомами). Ядро гранулоцитов обычно дольчатое (сегментированное), однако сравнительно немногочисленные менее зрелые их формы, циркулирующие в крови, имеют палочковидное ядро.
Агранулоциты (незернистые лейкоциты) содержат в цитоплазме лишь неспецифические (азурофильные) гранулы; специфические гранулы отсутствуют. Их ядро обычно имеет округлую или бобовидную форму. К агранулоцитам относятся моноциты и лимфоциты.
Лейкоцитарная формула. При проведении клинического анализа крови на ее мазках осуществляется дифференциальный подсчет относительного содержания лейкоцитов отдельных видов. Результаты такого подсчета регистрируются в табличной форме в виде так называемой лейкоцитарной формулы, в которой содержание клеток каждого вида представлено в процентах по отношению к общему количеству лейкоцитов, принятому за 100%:
Б |
Э |
|
Нейтрофилы |
|
Л |
Мон |
||
М |
Ю |
П |
С |
|||||
|
|
|
|
|||||
0.5-1 |
2-5 |
- |
0.5 |
3-5 |
60-65 |
20-35 |
6-8 |
|
Примечание: Б - базофилы, Э - эозинофилы, М - миелоциты, Ю - юные (метамиелоциты), Л - лимфоциты, Мон - моноциты.
НЕЙТРОФИЛЬНЫЕ ГРАНУЛОЦИТЫ
Нейтрофильные гранулоциты (нейтрофилы) - наиболее распространенный вид лейкоцитов и гранулоцитов. Они попадают в кровь из красного костного мозга, циркулируют в ней около 6-10 ч, частично располагаясь в пристеночном (близком к эндотелию), или маргинальном пуле, содержащем до 50% всех нейтрофилов крови. После циркуляции они мигрируют из крови в ткани, где функционируют от нескольких часов до 1-2 сут. (по некоторым данным, до 5-8 сут.). Они могут разрушаться значительно быстрее в очаге воспаления или в результате выхода на поверхность слизистых оболочек.
- 185 -
За сутки костный мозг взрослого человека выделяет в кровоток около 1011 нейтрофилов, столько же гибнет в тканях (преимущественно в слизистых оболочках и коже). Полагают, что существенная часть этих клеток (как и других гранулоцитов) в физиологических условиях погибает механизмом апоптоза без выделения цитотоксических продуктов их распада в окружающие ткани.
Функции нейтрофильных гранулоцитов:
1)Уничтожение микроорганизмов - возбудителей инфекций - основная функция нейтрофилов, отчего они считаются главными клеточными элементами неспецифической защиты организма. В связи со способностью к захвату (фагоцитозу) и уничтожению микробов И.И.Мечников назвал нейтрофилы микрофагами (в отличие от другой разновидности фагоцитов - макрофагов, поглощающих более крупные частицы). Нейтрофилы могут обеспечивать уничтожение микроорганизмов и без их поглощения - внеклеточно нефагоцитарными механизмами.
2)Разрушение и переваривание поврежденных клеток и тканей. Наиболее активно осуществляется на ранних сроках, так как нейтрофилы обычно первыми прибывают в очаг повреждения. Позднее эту функцию берут на себя макрофаги.
3)Участие в регуляции деятельности других клеток - осуществляется благодаря недавно установленной способности нейтрофилов к выработке ряда цитокинов, которая может резко усиливаться при стимуляции. Данная функция указывает на участие этих клеток не только в неспецифических, но и в специфических (иммунных) защитных реакциях.
Содержание нейтрофилов в крови взрослого в норме составляет:
относительное - 65-75% (от общего числа лейкоцитов), абсолютное - 3000-7000 клеток/мкл. Вследствие расположения около половины нейтрофилов в маргинальном (краевом) пуле их реальная абсолютная концентрация, как предполагают, примерно в два раза выше, чем определяемая при анализе крови.
Содержание нейтрофилов в крови ребенка меняется в зависимости от его возраста. Оно такое же, как у взрослого, непосредственно после рождения, затем оно падает и в период с 3-6 дней до 4-5 лет остается сниженным (до минимальных величин порядка 25%, типичных для первых двух лет жизни). После указанного периода оно возрастает, достигая уровня, характерного для взрослого, ко времени полового созревания.
- 186 -
Нейтропения - снижение содержания нейтрофилов в крови - обычно является следствием угнетения костного мозга в результате его аутоиммунного, токсического, лучевого или инфекционного поражения. При снижении концентрации нейтрофилов до 1000 клеток/мкл крови существенной опасности для здоровья обычно не возникает, однако при падении этого показателя до 500 клеток/мкл и ниже неизбежно развиваются тяжелые рецидивирующие инфекционные поражения.
Нейтрофилия - повышение содержания нейтрофилов в крови - возникает в результате усиленного выброса этих клеток из костного мозга при их значительном разрушении в ходе острого воспалительного (обычно инфекционного) процесса. Нейтрофилия при этом обычно сочетается с лейкоцитозом, а ее выраженность, как правило, пропорциональна активности воспалительного процесса. Умеренная нейтрофилия наблюдается также при физической нагрузке или эмоциональном стрессе, однако при этом она связана не с увеличением числа клеток в крови, а с их перераспределением - переходом части нейтрофилов из маргинального пула в общий (центральный) кровоток.
Размеры нейтрофильных гранулоцитов на мазках варьируют в пределах 10-15 мкм и примерно в 1.5 раза превышают размеры эритроцитов.
Плазмолемма нейтрофильных гранулоцитов обеспечивает разнообразные процессы, связанные с поддержанием жизнедеятельности и функциональной активности этих клеток. Она воспринимает различные сигналы, участвует в распознавании других клеток и компонентов межклеточного вещества (рецепторная функция), формировании многочисленных выпячиваний различной формы (в частности, связанных с движением клетки и фагоцитозом), транспорте веществ, процессах эндо- и экзоцитоза (в частности, дегрануляции). На плазмолемме находятся рецепторы адгезивных веществ, цитокинов, колониестимулирующих факторов (КСФ), медиаторов воспаления, иммуноглобулинов класса G, (IgG) СЗb-компонента комплемента, некоторых микробных продуктов.
Ядро нейтрофильных гранулоцитов имеет неодинаковое строение в клетках разyой степени зрелости. На основании строения ядра различают сегментоядерные, палочкоядерные и юные нейтрофильные гранулоциты.
Сегментоядерные нейтрофильные гранулоциты - наиболее зрелые,
составляют основную часть нейтрофилов (60-65% общего числа
- 187 -
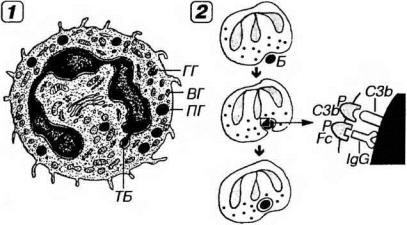
лейкоцитов). Для них характерно дольчатое ядро, которое представлено 2-5 (наиболее часто - 3-4) сегментами (рис. 7-8, см. также рис. 7-1), соединенными узкими нитевидными перетяжками (истончаются при созревании клетки). Оно интенсивно окрашено (преобладает гетерохроматин), что указывает на сравнительно низкую активность синтетических процессов в клетке. У женщин не менее 3% этих клеток содержат хорошо выявляемый дополнительный мелкий придаток ядра в виде барабанной палочки, который представляет собой неактивную X хромосому (половой хроматин, тельце Барра).
Рис. 7-8. Сегментоядерный нейтрофильный гранулоцит. 1 - ультраструктурная организация: ПГ - первичные (азурофильные) гранулы, ВГ - вторичные (нейтрофильные) гранулы, ГГ - гранулы гликогена. ТБ - тельце Барра, 2 - фагоцитоз бактерии (Б) нейтрофилом: последовательные стадии от адгезии нейтрофила к микробной клетке до ее захвата с формированием фагосомы. Активность поглощения бактерии резко возрастает при ее опсонизации вследствие взаимодействия покрывающих ее IgG и СЗb-компонента комплемента с рецепторами (Р) Fc-фрагмента IgG и С3b на плазмолемме нейтрофила.
Палочкоядерные нейтрофильные гранулоциты - более молодые клетки,
сравнительно немногочисленны (составляют 3-5% общего числа лейкоцитов). Их ядро (в форме палочки, подковы или изогнутой колбаски) не сегментировано или содержит лишь намечающиеся перетяжки, которые углубляются по мере созревания клеток (см. рис. 7-1). В части палочкоядерных нейтрофилов ядро содержит меньше гетерохроматина, чем в сегментоядерных. Относительное содержание палочкоядерных форм является показателем скорости поступления нейтрофилов в кровоток. Оно обычно повышается при нейтрофилии, сочета-
- 188 -
ясь в выраженных случаях с нарастанием числа юных нейтрофилов (метамиелоцитов), что оценивается как "сдвиг влево" на гемограмме (юные <- палочкоядерные <— сегментоядерные), в которой молодые формы клеток записываются левее более зрелых (см. выше). Выраженный сдвиг влево отмечается у новорожденых в течение 1-й нед. жизни.
Юные нейтрофильные гранулоциты (метамиелоциты) - наиболее молодые клетки нейтрофильного ряда среди тех, что в норме встречаются в крови. Они обнаруживаются в чрезвычайно малом количестве (до 0.5% обшего числа лейкоцитов). Их ядро имеет бобовидную форму и светлее, чем у палочко и сегментоядерных клеток.
Цитоплазма нейтрофильных гранулоцитов на светооптическом уровне слабооксифильна. При электронно-микроскопическом исследовании в ней выявляются немногочисленные органеллы: отдельные элементы грЭПС, митохондрии, свободные рибосомы, мелкий комплекс Гольджи, пентриоли. Из включений преобладают гранулы гликогена.
Цитоскелет нейтрофильных гранулоцитов представлен небольшим числом (12-20/клетку) микротрубочек, умеренно развитыми виментиновыми промежуточными филаментами и многочисленными актиновыми микрофиламентами, расположенными преимущественно в периферической части цитоплазмы, образующей псевдоподии и свободной от других органелл и включений. В покоящихся нейтрофилах менее половины актина находится в виде полимера (Р-актина), основная же его часть представлена неполимеризованным глобулярным С-актином. При стимуляции клетки уже в течение нескольких секунд до 90% имеющегося актина полимеризуется с образованием филаментов в подмембранной зоне, в особенности, в участке фагоцитоза.
Цитоплазматические гранулы нейтрофилов сравнительно многочисленны (по 50-200 в каждой клетке) и разделяются на три типа: первичные, вторичные и третичные. Помимо гранул выявлены также мембранные секреторные пузырьки. Согласно современным представлениям, гранулы нейтрофилов не являются сугубо изолированными образованиями, а образуют единую функциональную систему с самостоятельными, но частично перекрывающимися функциями и составом компонентов (в частности, во всех видах гранул содержится лизоцим).
- 189 -
1. Первичные (азурофилъные, или неспецифические) гранулы названы так потому, что появляются первыми в ходе развития (на стадии промиелоцита - см. главу 9). В зрелых клетках они составляют
лишь 10-30% общего числа гранул, окрашиваются азуром в розово-фиолетовый цвет и не являются специфическими для нейтрофилов, поскольку встречаются и в лейкоцитах других типов. Эти гранулы имеют самые крупные размеры (диаметр 400-800 нм, в среднем около 500 нм) и соответствуют зернистости, выявляемой на светооптическом уровне. Они имеют вид округлых или овальных мембранных пузырьков с электронно-плотным содержимым и часто рассматриваются как лизосомы. В них, однако, имеется большой набор антимикробных веществ, что не характерно для обычных лизосом.
Вещества, содержащиеся в первичных гранулах - лизоцим, миелопероксидаза, нейтральные протеиназы, кислые гидролазы, дефензины (на которые приходятся 30-50% белка гранул), катионные анти-микробные белки, бактерицидный белок, увеличивающий проницаемость (BPI-белок - от англ.
Bactericidal Permeability Increasing), - обладают высокой микробицидной активностью. Ферменты этих гранул активны преимущественно в кислой среде
иобеспечивают внутриклеточное уничтожение микробов.
2.Вторичные (специфические) гранулы появляются позднее первичных в процессе развитая (в конце стадии промиелоцита и особенно активно на стадии миелоцита - см. главу 9) и становятся все более многочисленными при созревании нейтрофилов; в зрелых клетках они составляют 80-90% общего числа гранул. Они плохо выявляются под световым микроскопом, так как имеют мелкие размеры (диаметр - 100-300 им; в среднем - 200 нм - на границе разрешения светового микроскопа). При электронной микроскопии они имеют вид мембранных пузырьков округлой, овальной или гантелевидной формы с зернистым содержимым сравнительно низкой плотности.
Вещества, содержащиеся во вторичных гранулах (лизоцим, лактоферрин, щелочная фосфатаза, коллагеназа, активатор ппазминогена, частично - катионные белки) участвуют во внутриклеточном разрушении микробов, а также секретаруются в межклеточное вещество, где они играют роль в мобилизации медиаторов воспалительной реакции и активации системы комплемента. В этих гранулах содержатся также адгезивные белки.
3. Третичные (желатинозные) гранулы нейтрофильных гранулоцитов оцисаны недавно и изучены неполностью. По размерам и морфологическим характеристикам они сходны со специфическими гранулами, но отличаются от них по химическому составу. Главными компонентами содержимого этих гранул являются желатиназа (обнаружена
- 190 -