

Тетрагидрофолиевая кислота и ее роль в синтезе и переносе одноуглеродных радикалов.
В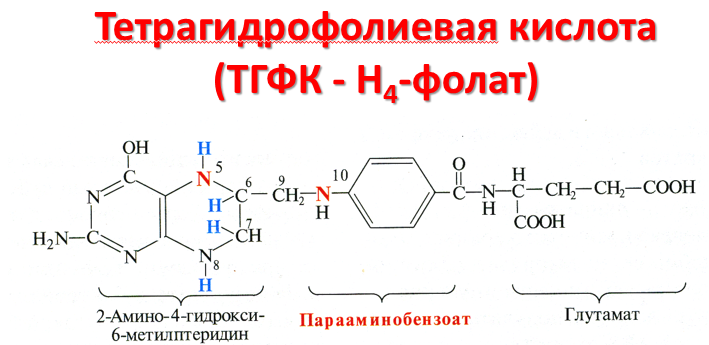 превращениях серина и глицина главную
роль играют ферменты, коферментами
которых служат производные фолиевой
кислоты. Этот витамин широко распространён
в животных и растительных пищевых
продуктах. Молекула фолиевой кислоты
(фолата) состоит из 3 частей: птеринового
производного, парааминобензойной и
глутаминовой кислот.
превращениях серина и глицина главную
роль играют ферменты, коферментами
которых служат производные фолиевой
кислоты. Этот витамин широко распространён
в животных и растительных пищевых
продуктах. Молекула фолиевой кислоты
(фолата) состоит из 3 частей: птеринового
производного, парааминобензойной и
глутаминовой кислот.
Г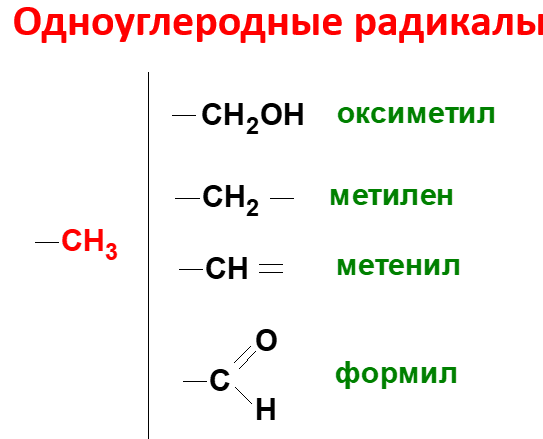 омоцистеин
может снова превращаться в метионин
под действием гомоцистеинметил-транс-феразы.
Донором метильной группы в этом случае
служит N5-метил-Н4-фолат:
омоцистеин
может снова превращаться в метионин
под действием гомоцистеинметил-транс-феразы.
Донором метильной группы в этом случае
служит N5-метил-Н4-фолат:
Образование и использование одноуглеродных фрагментов.Особое значение реакций катаболизма серина и глицина заключается в том, что они сопровождаются образованием одноуглеродного метиленового фрагмента (-СН2-). Метиленовая группа в молекуле метилен- Н4-фолата может превращаться в другие одноуглеродные группы (фрагменты): метенильную (-СН=), формильную (-НС=О), метильную (-СН3) и формиминогруппу (-CH=NH
В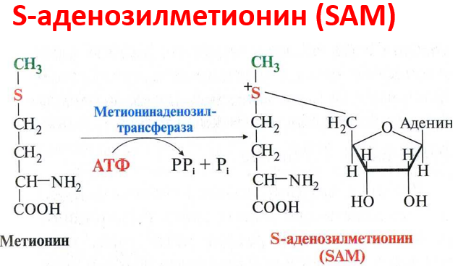 се
образующиеся производные Н4-фолата
играют роль промежуточных переносчиков
и служат донорами одноуглеродных
фрагментов при синтезе некоторых
соединений: пуриновых оснований и
тимидиловой кислоты (необходимых для
синтеза ДНК и РНК), регенерации метионина,
синтезе различных формиминопроизводных
(формиминоглицина и т.д.)
се
образующиеся производные Н4-фолата
играют роль промежуточных переносчиков
и служат донорами одноуглеродных
фрагментов при синтезе некоторых
соединений: пуриновых оснований и
тимидиловой кислоты (необходимых для
синтеза ДНК и РНК), регенерации метионина,
синтезе различных формиминопроизводных
(формиминоглицина и т.д.)
Обмен фенилаланина и тирозина. Пути превращения в норме.
Метаболизм фенилаланина
Основное количество фенилаланина расходуется по 2 путям:
• включается в белки;
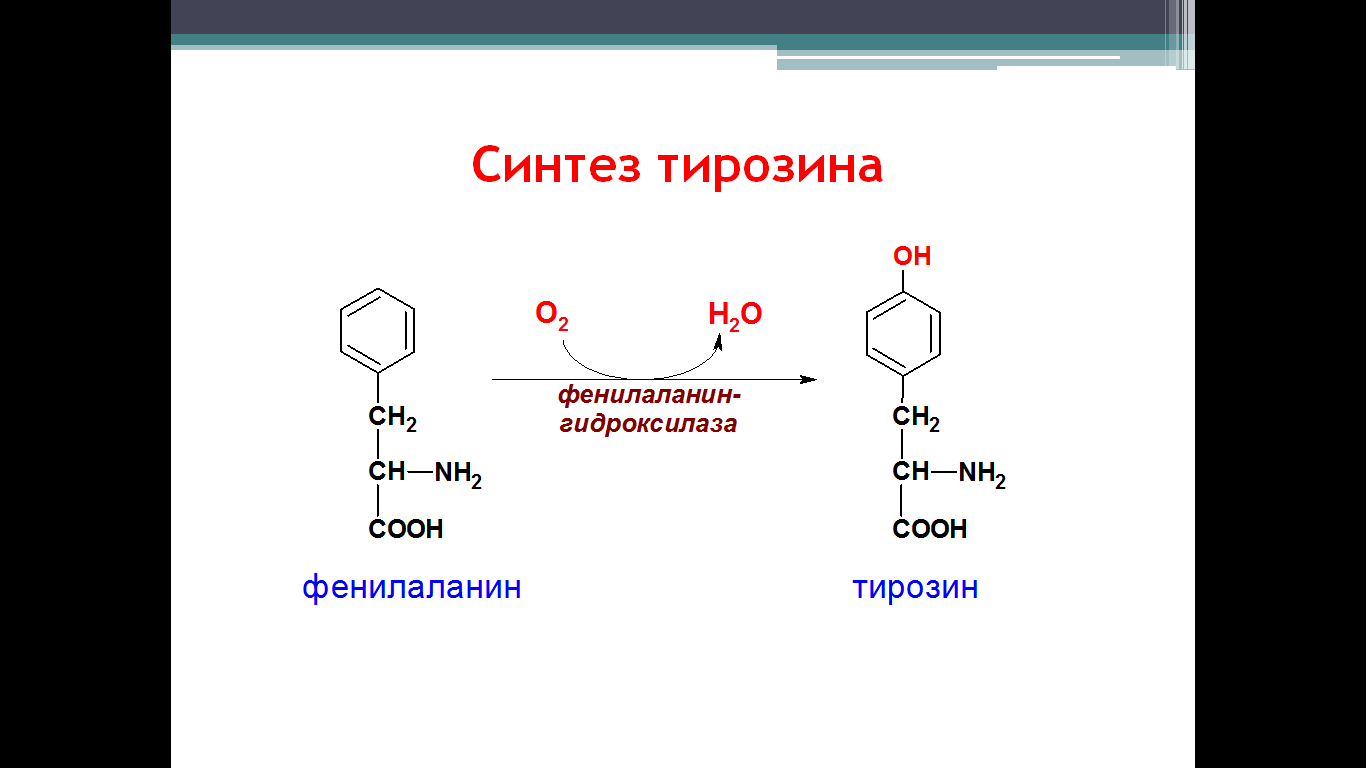
• превращается в тирозин. Превращение фенилаланина в тирозин прежде
всего
необходимо для удаления избытка
фени-лаланина, так как высокие концентрации
его токсичны для клеток. Образование
тирозина не имеет большого значения,
так как недостатка этой аминокислоты
в клетках практически не бывает.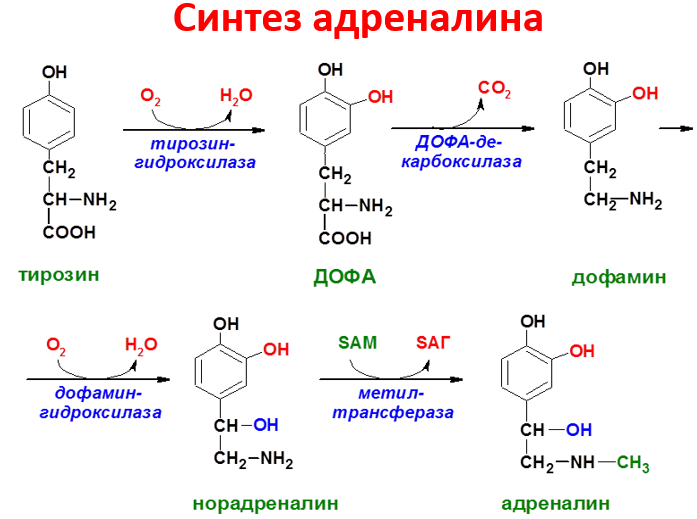
Фенилаланин - незаменимая аминокислота, так как в клетках животных не синтезируется её бензольное кольцо.
Тирозин - условно заменимая аминокислота, поскольку образуется из фенилаланина. Содержание этих аминокислот в пищевых белках (в том числе и растительных) достаточно велико. Фенилаланин и тирозин используются для синтеза многих биологически ктивных соединений. В разных тканях метаболизм этих аминокислот происходит по-разному
М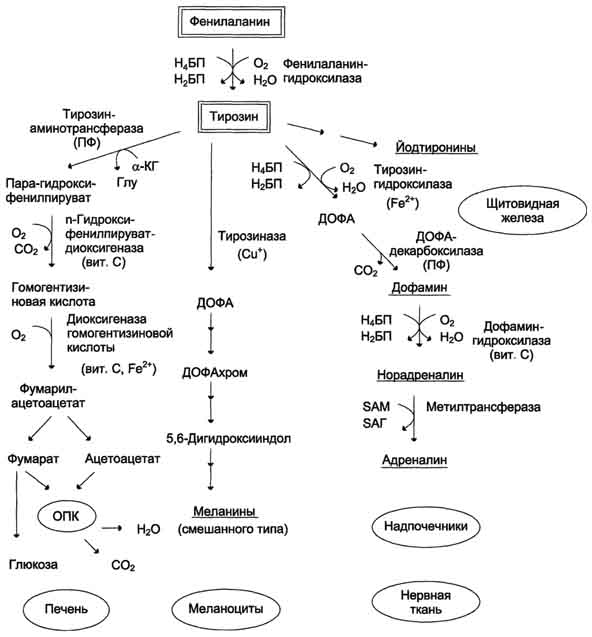 етаболизм
феиилаланина
етаболизм
феиилаланина
Основное количество фенилаланина расходуется по 2 путям:
включается в белки;
превращается в тирозин.
Превращение фенилаланина в тирозин прежде всего необходимо для удаления избытка фенилаланина, так как высокие концентрации его токсичны для клеток. Образование тирозина не имеет большого значения, так как недостатка этой аминокислоты в клетках практически не бывает. Основной путь метаболизма фенилаланина начинается с его гидроксилирования, в результате чего образуется тирозин. Эта реакция катализируется специфической монооксиге-назой - фенилаланингидр(жсилазой, кофермен-том которой служит тетрагидробиоптерин (Н4БП). Активность фермента зависит также от наличия Fe2+. Реакция необратима. Н4БП в результате реакции окисляется в дигидробиоптерин (Н2БП). Регенерация последнего происходит при участии дигидроптеридинредуктазы с использованием NADPH + H+. Обмен тирозина значительно сложнее, чем обмен фенилаланина. Кроме использования в синтезе белков, тирозин в разных тканях выступает предшественником таких соединений, как катехоламины, тироксин, меланины, и ка-таболизируется до СО2 и Н2О.
Катаболизм тирозина в печени
В печени происходит катаболизм тирозина до конечных продуктов. Специфический путь катаболизма включает несколько ферментативных реакций, завершающихся образованием фумарата и ацетоацетата Трансаминирование тирозина с ос-кетоглутаратом катализирует тирозинаминотрансфе-раза(кофермент ПФ) - индуцируемый фермент печени млекопитающих. В результате образуется п-гидроксифенилпируват.
В реакции окисления п-гидроксифенилпирувата в гомогентизиновую кислоту происходит декарбоксилирование, гидроксилирование ароматического кольца и миграция боковой цепи. Реакцию катализирует фермент n-гидроксифенилпируватдиоксигеназа, кофакторами которого выступают витамин С и Fe2+.
Превращение гомогентизиновой кислоты в фумарилацетоацетат сопровождается расщеплением ароматического кольца. Эта реакция катализируется диоксигеназой гомогентизиновой кислоты, в качестве кофермента содержащей Fe2+.
Обмен фенилаланина и тирозина связан со значительным количеством реакций гидроксилирования, которые катализируют оксигеназы. Ферменты оксигеназы (гидроксилазы) используют молекулу О2 и кофермент-донор водорода (чаще - Н4БП). Для катализа оксигеназам необходимы кофакторы - Fe2+ или гем (для некоторых - Сu+), а для многих ещё и витамин С. Оксигеназы делят на 2 группы:
Монооксигеназы - один атом О2 присоединяют к продукту реакции, другой используют для образования Н2О;
Диоксигеназы - оба атома О2 используют для образования продукта реакции.
Почти все процессы расщепления ароматических колец в биологических системах катализируются диоксигеназами, подклассом ферментов, открытым японским биохимиком Осами Хайяши. В результате разрыва бензольного кольца образуется малеилацетоацетат, который в процессе цис- и транс-изомеризации превращается в фумарилацетоацетат.
Гидролиз фумарилацетоацетата при действии фумарилацетоацетатгидролазы приводит к образованию фумарата и ацетоацетата. Фумарат может окисляться до СО2 и Н2О или использоваться для глюконеогенеза. Ацетоацетат - кетоновое тело, окисляемое до конечных продуктов с выделением энергии.
Превращение тирозина в меланоцитах. В пигментных клетках (меланоцитах) тирозин выступает предшественником тёмных пигментов - меланинов. Среди них преобладают 2 типа: эумеланины и феомеланины. Эумеланины (чёрного и коричневого цвета) - нерастворимые высокомолекулярные гетерополимеры 5,6-дигидроксииндола и некоторых его предшественников. Феомеланины - жёлтые или красновато-коричневые полимеры, растворимые в разбавленных щелочах. Находятся они, в основном, в составе волос. Меланины присутствуют в сетчатке глаз. Цвет кожи зависит от распределения меланоцитов и количества в них разных типов меланинов.
Превращение тирозина в щитовидной железе В щитовидной железе синтезируются и выделяются гормоны йодтиронины: тироксин (тетрайодтиронин) и трийодтиронин. Эти гормоны представляют собой йодированные остатки тирозина, которые попадают в клетки щитовидной железы через базальную мембрану
Превращения тирозина в надпочечниках и нервной ткани (синтез катехоламинов)В мозговом веществе надпочечников и нервной ткани тирозин является предшественником катехоламинов (дофамина, норадреналина и адреналина) При образовании катехоламинов, которое происходит в нервной ткани и надпочечниках, и меланина в меланоцитах промежуточным продуктом служит диоксифенилаланин (ДОФА) . Однако гидроксилирование тирозина в клетках различных типов катализируется различными ферментами:
Тирозиназа в меланоцитах является Сu+-зависимым ферментом (см. выше).
Тирозингидроксилаза в надпочечниках и катехоламинергических нейронах не нуждается в ионах меди. Это - Fе2+-зависимый фермент, аналогично фенилаланингидроксилазе в качестве кофермента использующий Н4БП.
Физиологическая роль тирозингидроксилазы чрезвычайно велика, так как этот фермент является регуляторным и определяет скорость синтеза катехоламинов.
Активность тирозингидроксилазы значительно изменяется в результате:
Аллостерической регуляции (ингибитор - норадреналин);
Фосфорилирования/дефосфорилирования: в результате фосфорилирования с участием протеинкиназы А снижаются Кm для кофермента Н4БП и сродство фермента к норадреналину, в результате чего происходит активация тирозингидроксилазы.
Количество фермента регулируется на уровне транскрипции.
ДОФА-декарбоксилаза (кофермент - ПФ) катализирует образование дофамина, который при участии дофамингидроксилазы (монооксигеназы) превращается в норадреналин. Для функционирования фермента необходимы ионы Сu+, витамин С и тетрагидробиоптерин.
В мозговом веществе надпочечников фенилэтаноламин-N-метилтрансфераза катализирует метилирование норадреналина, в результате чего образуется адреналин. Источником метальной группы служит SАМ.
Дофамин и норадреналин служат медиаторами в синаптической передаче нервных импульсов, а адреналин - гормон широкого спектра действия, регулирующий энергетический обмен. Одна из функций катехоламинов - регуляция деятельности ССС