

6.3 Электропроводность клеток и тканей для переменного тока
Биологические системы способны накапливать электрические заряды при прохождении через них переменного тока, поэтому для описания их свойств наряду с сопротивлением используют понятия электрической емкости (С):
С = Δq/Δφ, (6.9)
где: Δq – изменение заряда; Δφ – изменение потенциала.
Электрическая емкость объекта при равных условиях его геометрии определяется диэлектрической проницаемостью, которая описывает все явления поляризации в биообъекте:
ε · S
С = ————, (6.10)
4 π · d
где: С – электрическая емкость; S – площадь поверхности (или пластин конденсатора); d – расстояние между поверхностями (или пластинами); ε – диэлектрическая проницаемость.
Емкость биообъекта определяется поляризационной емкостью, которая возникает в момент прохождения электрического тока. Она отражает отношение изменения заряда объекта к изменению его потенциала при прохождении через биообъект переменного тока. Изменение заряда за время t определяется по формуле:
Δq = о∫t I · dt; (6.11)
изменение потенциала:
Δφ = R · (IO – It); (6.12)
о∫t I · dt
тогда поляризационная емкость СР = —————, (6.13)
R · (IO – It)
где: IO и It – начальное и конечное значение силы тока; I – мгновенный ток; R – сопротивление.
К поляризационной емкости биообъекта присоединяется большая по величине статическая емкость мембраны (около 1 мкф/см2). Величина поляризационной емкости (уравнение 6.13) зависит от времени действия внешнего поля и может при низкой частоте превышать величину статической емкости.
При высокой частоте тока (более 10 кГц) статическая емкость на несколько порядков превышает поляризационную емкость. Поскольку данные емкостные части систем соединены последовательно, то при высокой частоте общая величина емкости определяется меньшей составляющей, то есть поляризационной емкостью. Поскольку биообъекты обладают проводимостью и емкостью, они характеризуются “активным” и “реактивным” сопротивлением. “Реактивное” емкостное сопротивление RX определяется по формуле:
RX = 1/ω · C, (6.14)
где: ω – частота тока. Суммарное сопротивление биообъектов называют импеданс.
Электрическая модель биообъекта может быть представлена в виде эквивалентных схем с последовательным и параллельным сопротивлением С и R (рис.6.4):
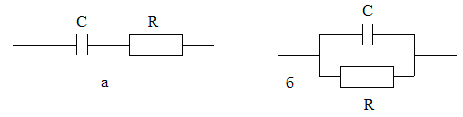
Рис.6.4. Последовательное (а) и параллельное (б) соединение емкостного и обычного сопротивления.
Для последовательного соединения С и R импеданс определяют по формуле:
Z = R – i · 1/ω · C или по формуле: Z2 = R2 + 1/(ω2 · C2); (6.15)
для параллельного – по формуле:
1/Z = 1/R + i · ω · C,
где: Z – импеданс; i – мнимая единица, то есть (-1)1/2. Импеданс биообъектов зависит от частоты тока: при увеличении частоты реактивная составляющая импеданса уменьшается. Такая зависимость является одной из причин дисперсии импеданса.
При изучении частотных зависимостей сопротивления и емкости биообъектов было обнаружено три области дисперсии (рис.6.5).

Рис.6.5. Три области дисперсии импеданса.
1.α – Дисперсия занимает область низких частот звукового диапазона, примерно до 1 кГц. В этой области уменьшение диэлектрической проницаемости биосистем обусловлено уменьшением эффекта поляризации поверхности клеток, поскольку электрический ток с частотой до 1 кГц протекает только по межклеточным структурам. Сопротивление самих клеток для токов такой частоты очень велико.
Явления поляризации, происходящие на биологических поверхностях, аналогичны явлениям поляризации на электродах. В обоих случаях поляризация возникает за счет перераспределения ионов в двойном электрическом слое. В таком случае поверхностная поляризация сводится к электрической поляризации.
По мере увеличения частоты тока эффект поверхностной поляризации уменьшается. Это проявляется как снижение диэлектрической проницаемости и сопротивления клеток.
2.β – Дисперсия занимает область частот 1,0 – 104 кГц. В данной области электрические свойства биообъектов наиболее полно описываются с помощью мактроструктурной поляризации. Она вызвана наличием не только клеточной мембраны, но и мембранными структурами в цитоплазме клетки.
Если в данном случае применить электрохимическую теорию, то сущность состоит в перераспределении ионов на границе макроструктурного объема. В данном случае протекает электролитическая поляризация границы раздела фаз.
На достаточно высоких частотах, порядка нескольких мегагерц, поляризация границ раздела, то есть мембран, будет отсутствовать.
3.γ – Дисперсия наблюдается на частотах свыше 103 мГц. Уменьшение диэлектрической проницаемости в данном диапазоне обусловлено ослаблением эффекта поляризации, вызванной диполями воды. Величина γ – дисперсии будет зависеть от содержания свободной воды в исследуемой ткани.
В области сверхвысоких частот (> 104 мГц) эффект поляризации, обусловленной диполями воды, будет отсутствовать. Диэлектрическая проницаемость будет иметь небольшие значения, которые определяются ионной и электронной поляризацией с самым небольшим временем релаксации.
На три основные дисперсионные явления (α –, β –, γ –), обусловленные поверхностной поляризацией, макроструктурной и дипольной поляризацией воды, накладываются дополнительные эффекты:
1.Поляризация белков и других органических молекул, обладающих дипольными моментами. Дисперсия данного вида поляризации происходит при частотах в несколько мегагерц.
2.Поляризация гидратных оболочек макромолекул. Дисперсия наблюдается в диапазоне частот 10,0 – 100,0 мГц.
3.Поляризация связанных полярных групп макромолекул. Область дисперсии данного вида поляризации наблюдается в диапазоне 100,0 – 1000,0 мГц.