

С
1 — кора
великого мозку; 2 —
довгастий мозок; 3
— спинний мозок; 4
— провідний аферентний
шлях; 5
— колатералі від провідних шляхів до
нейронів сітчастого утвору; 6
— сітчастий утвір; 7
— ядра таламуса; 8 —
висхідна активаційна
система; 9 —
низхідна активаційна система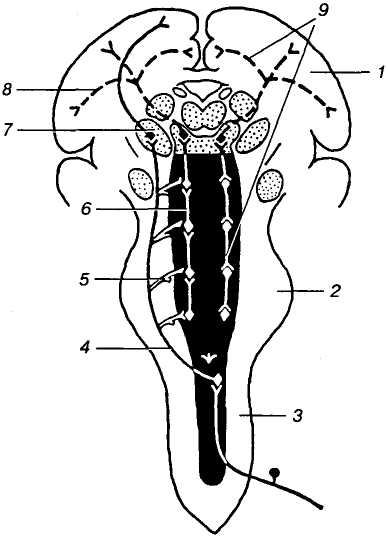
Мал. 18. Сітчастий утвір:
Нейрони сітчастого утвору мають по 1-6 довгих розгалужених дендритів і аксон, дві гілки якого утворюють: одна — еферентні сітчастоспинномозкові шляхи, а інша — висхідні шляхи. До нейронів сітчастого утвору підходить велика кількість колатералей від усіх специфічних висхідних шляхів, які проходять через стовбур мозку, а також від низхідних шляхів, що йдуть від вищих відділів мозку, в тому числі від кори великого мозку й мозочка. Отже, за своєю нейронною організацією і зв'язками сітчастий утвір поєднує впливи різних мозкових структур.
Майже всі нейрони сітчастого утвору є полісенсорними, тобто можуть реагувати на подразнення кількох модальностей, наприклад, на подразнення шкіри та дію світла, на больові й звукові подразнення. Ці нейрони мають також високу хімічну чутливість і тому легко пригнічуються деякими лікарськими речовинами, особливо барбітуратами, які повністю блокують їх. Ця обставина робить неможливим дослідження сітчастого утвору під наркозом.
Функція сітчастого утвору зумовлена особливостями її нейронів. Однією з таких особливостей є здатність до підтримання стійкої тонічної активності. Практично всі нейрони безперервно генерують імпульси з частотою 5—10 за 1 с. Різні аферентні сигнали підсумовуються з цією фоновою активністю, викликаючи в одних клітинах її збільшення, а в інших, навпаки, гальмування.
Тонічна активність сітчастого утвору. Постійна фонова активність нейронів сітчастого утвору пояснюється, з одного боку, їх високою хімічною чутливістю до впливу гуморальних чинників, які постійно деполяризують клітинну мембрану і збуджують нейрони. З іншого боку, це визначається особливостями аферентних зв'язків сітчастих нейронів, на яких конвертують колатералі від величезної кількості різних чутливих волокон, що зумовлює постійне подразнення сітчастих нейронів імпульсами чутливих систем організму. Внаслідок цього сітчасті нейрони перебувають у стані постійної активності, що й зумовлює безперервний тонічний вплив сітчастого утвору на розміщені вище і нижче структури мозку.
Дихальний і судиноруховий центри розміщені в межах сітчастого утвору довгастого мозку. Нейрони цих центрів займають порівняно невелику ділянку, розміщену в його задньому відділі. Функція решти сітчастого утвору довгий час була невідомою.
Висхідні впливи сітчастого утвору. У 1949 р. Г. Мегун і Дж. Моруцці встановили, що подразнення певної частини сітчастого утвору має стимулювальний вплив на функцію кори великого мозку, подразнення інших її ділянок — гальмівний. Так, якщо тварина перебувала у стані сну, то після подразнення збуджувальної частини сітчастого утвору вона прокидалася і поводилась активно. Одночасно у мозку виникала реакція десинхронізації, яка охоплювала значні ділянки кори, що свідчить про генералізований характер висхідних активаційних впливів сітчастого утвору.
Реакція десинхронізації не єдиний прояв впливу сітчастого утвору на функцію кори великого мозку. Відомо, що при будь-якому аферентному збудженні у відповідних проекційних полях кори з'являється своєрідна електрична реакція, яка дістала назву первинної відповіді.
Невдовзі за первинною відповіддю виникають додаткові коливання електричного потенціалу, які мають більший латентний період і генералізований характер. Це реакції вторинної відповіді. Спеціальні дослідження показали, що вторинні відповіді є електричним проявом надходження аферентного збудження до кори великого мозку за допомогою сітчасто-кіркових волокон.
Отже, при аферентному збудженні імпульсація через стовбур головного мозку і таламус потрапляє до кори великого мозку, що реєструється у вигляді первинної відповіді у проекційній зоні кори. Ця реакція має невеликий латентний період. Одночасно частина аферентної імпульсації по колатералях відгалужується до сітчастого утвору, активуючи його. Висхідними сітчасто-кірковими шляхами імпульсація надходить до кори, але вже у вигляді вторинної (затриманої) відповіді, яка виникає з довшим латентним періодом. Ця реакція охоплює не лише проекційну зону, а й сусідні ділянки кори, викликаючи в них стан підвищеної збудливості. Сумарну електричну відповідь на аферентне збудження називають викликаним потенціалом.
Сітчастий утвір може виявляти й гальмівний вплив на ділянки ЦНС, розміщені вище, хоча організація гальмівної висхідної системи з'ясована не до кінця.
Низхідні впливи сітчастого утвору. Полегшувальні й гальмівні імпульси від сітчастого утвору проводяться також до спинного мозку сітчасто-спинномозковим шляхом (див. мал. 18), регулюючи його функцію. Так, подразнення сітчастого гігантоклітинного ядра у довгастому мозку зумовлює пригнічення рефлекторної активності спинного мозку, причому це пригнічення є дуже глибоким і тривалим, навіть у тому випадку, якщо подразнення цього ядра спричинене лише кількома стимулами. Це дифузне неспецифічне гальмування охоплює майже всі рефлекторні дуги і ті нейрони, які беруть участь у передачі висхідної інформації, що призводить не лише до пригнічення рефлекторної діяльності спинного мозку, а й викликає часткову сенсорну депривацію кори великого мозку.
У 1862 p. I. M. Сеченов виявив факт пригнічення спінальних рефлексів при подразненні стовбура головного мозку. Це було відкриттям центрального гальмування і одночасно сітчасто-спинномозкової системи. Проте механізм цього гальмівного ефекту вдалося розкрити лише майже через 100 років — у 50-ті роки XX ст. завдяки дослідженням Г. Мегуна, який показав, що локальне електричне подразнення гігантоклітинного ядра довгастого мозку викликає неспецифічне гальмування згинальних і розгинальних спинномозкових рефлексів. Механізм сєченівського гальмування виявився у тому, що збудження нейронів сітчастого утвору, яке поширюється через сітчасто-спинномозковий шлях, активує гальмівні вставні нейрони спинного мозку, аксони яких утворюють гальмівні синапси на α-мотонейронах. При цьому мембрана α-мотонейронів гіперполяризується, а їхня збудливість відповідно знижується.
Крім гальмівного сітчастий утвір може виявляти й полегшувальний вплив на функцію спинномозкових структур. Центр полегшувальної системи сітчастого утвору знаходиться у ділянці моста і середнього мозку.
Сітчастий утвір як прямо, так і опосередковано взаємодіє майже з усіма структурами і системами мозку, впливаючи на різноманітні функції. Одна з найголовніших — це функція неспецифічного підвищення збудливості, пов'язана з процесами активації та регуляції рівня притомності й уваги.
С
ЗАДНІЙ МОЗОК
Задній мозок представлений двома філогенетично і функціонально пов'язаними структурами — мостом і мозочком. У процесі еволюції мозочок відділився від решти мозку і перетворився на анатомічно відносно автономну структуру, пов'язану зі стовбуром мозку трьома парами ніжок. У зв'язку з цим будова і функції мозочка розглядаються, як правило, окремо.
Міст (вароліїв). Саме через міст проходять кірково-мозочкові шляхи. Міст розміщений між довгастим і середнім мозком, займає у людини передню частину заднього мозку і складається з двох структурно-функціональних утворів: основної частини і покриву моста.
В основній частині моста проходять кірково-спинномозкові шляхи, між якими розміщуються власні нервові волокна моста, що прямують до мозочка. З основи моста виходять нерви, які зв'язують його певні зони з корою великого мозку.
У ділянці покриву моста розміщені ядра сітчастого утвору, а також частково присінково-завиткового нерва. Якщо нейрони ядер завитки сприймають сигнали від рецепторів спірального органа завиткової протоки, а нейрони верхнього присінкового ядра — від рецепторів присінка і півколових каналів, то присереднє і бічне присінкові ядра стосуються контролю рухових функцій, їхні нейрони дають початок присінково-спинномозковому шляху, аксони якого досягають рухових нейронів спинного мозку.
У ділянці покриву також розміщені рухові ядра трійчастого, лицевого та язикоглоткового нервів. Вони іннервують м'язи лиця, глотки. До цієї самої групи функціонально входять і рухові ядра черепних нервів довгастого мозку, які іннервують м'язи шиї. Вони беруть участь у здійсненні рухових рефлексів, спрямованих на відновлення і збереження постави.
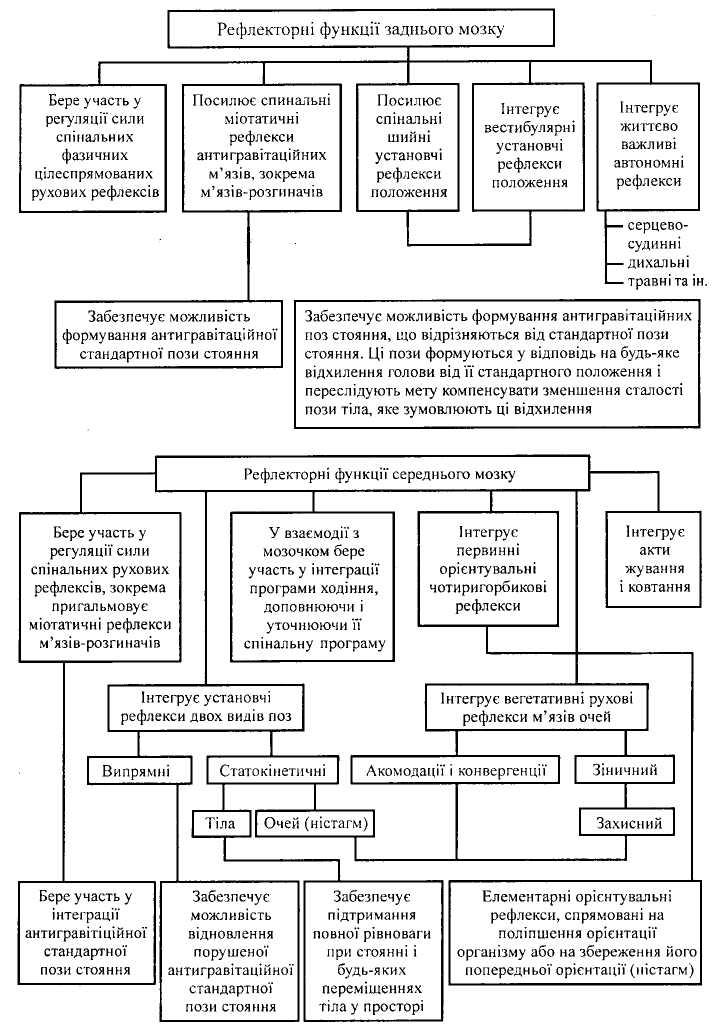
Р
Мал. 19.
Головні рефлекси заднього
мозку
До вегетативних належать рефлекси, що регулюють за допомогою парасимпатичних ядер лицевого і язикоглоткового нервів функцію слинних залоз. Крім того, ядра моста беруть участь у регуляції сльозо-і потовиділення, координації актів жування і ковтання, кліпання очима тощо. Парасимпатичне ядро блукаючого нерва входить до системи рефлекторної регуляції дихання і кровообігу.
С
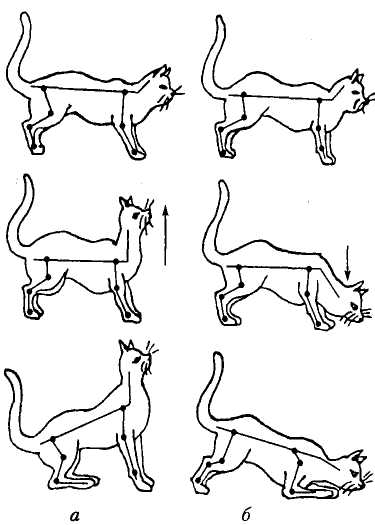
Мал. 20. Шийні тонічні рефлекси
у децереброваного кота з видаленими
присінками: а — пасивне
піднімання голови (показано стрілкою)
супроводжується підвищенням тонусу
розгиначів передніх кінцівок і зниженням
— задніх; б — пасивне
згинання голови зумовлює протилежний
ефект
Тонічні, або постуральні (фр. posture — положення тіла, поза), (статичні) рефлекси — це група рефлексів, у здійсненні яких беруть участь структури довгастого, заднього і середнього мозку. Вони спрямовані на підтримання певного положення тіла (пози) чи його частини у просторі. Залежно від джерела аферентних сигналів тонічні рефлекси поділяють на шийні і присінкові.
III и й н і тонічні рефлекси викликаються збудженням пропріорецепторів м'язів шиї. Вперше вони були описані Р. Магнусом у котів з перетином стовбура головного мозку. У чистому вигляді ці рефлекси можна спостерігати лише після руйнування присінкового апарату, який надає відповідну інформацію про положення голови у просторі.
Як показали досліди Р. Магнуса, закидання котові голови назад супроводжується зростанням тонусу м'язів — розгиначів передніх кінцівок з одночасним зниженням тонусу м'язів — розгиначів задніх кінцівок (мал. 20). Згинання голови викликає протилежні реакції, а повороти голови праворуч і ліворуч стосовно поздовжньої осі зумовлюють компенсаторне підвищення тонусу м'язів — розгиначів тих кінцівок, у бік яких повернено голову. Це рефлекси обертання за Р. Магнусом.
Ефекторною ланкою шийних рефлексів можуть бути і м'язи очного яблука, оскільки вони рефлекторно забезпечують компенсаторне встановлення очей зі зміною положення голови.
Усі шийні рефлекси є полісинаптичними. Імпульси від пропріорецепторів шийних м'язів проходять через клиноподібне ядро (Бурдаха) до відповідних центрів довгастого мозку, а звідти до рухових нейронів спинного мозку або рухових ядер відвідного нерва і викликають зміни положення тіла і тонусу м'язів очного яблука.
Присінкові (вестибулярні) рефлекси поділяють на статичні і статокінетичні. Статичні рефлекси пов'язані переважно зі збудженням рецепторів перетинчастого лабіринту присінка і забезпечують підтримання пози й рівноваги тіла при змінах його положення у просторі — рефлекси пози, рефлекси випрямлення. Наприклад, якщо кіт падає з висоти спиною донизу, то спочатку у нього завдяки присінковому рефлексу випрямлення відновлюється нормальне положення голови (мордою донизу). Ця зміна положення голови збуджує пропріорецептори шийних м'язів, спрацьовує шийний рефлекс випрямлення, внаслідок чого нормального положення набуває також тулуб.
Статокінетичні рефлекси спрямовані на підтримання пози у разі зміни швидкості руху. Ці рефлекси зумовлені збудженням рецепторів півколових каналів, яке виникає під час руху ендолімфи всередині каналів. Прискорення під час обертання тіла у горизонтальній площині збуджує рецептори горизонтального півколового каналу і викликає рефлекторну реакцію окорухового апарату — горизонтальний очний ністагм. Ця реакція полягає в тому, що в момент прискорення обертання очні яблука рухаються в бік, протилежний напрямку обертання. Досягнувши максимально можливого відхилення, очні яблука швидко переміщуються у напрямку руху, і в поле зору потрапляє інша точка простору.
Ністагм сприяє збереженню нормальної зорової орієнтації. Цю реакцію використовують також з діагностичною метою для перевірки функціонування статокінетичного аналізатора (присінкового апарату). Площина ністагму збігається з площиною обертання, ось чому крім горизонтального розрізняють також вертикальний, діагональний та коловий ністагм.
До статокінетичних рефлексів належать також рефлекси ліфта: при підійманні в ліфті з прискоренням кінцівки розгинаються, а при опусканні — згинаються.
Статичні і статокінетичпі рефлекси є у людини вже під час внутрішньоутробного розвитку. Після народження, в міру дозрівання і вдосконалення центральних механізмів координації нервово-м'язового апарату у дитини послідовно виникають рефлекторні реакції, які зумовлюють утримання голови, сидіння, стояння і ходіння. У подальшому відбувається вдосконалення процесів управління позами і рухами, зокрема розвивається кірковий контроль за функцією нервових центрів заднього мозку.
СЕРЕДНІЙ МОЗОК
Середній мозок — це порівняно невелика частина стовбура головного мозку, розміщена між заднім (мостом) і проміжним мозком.
Середній мозок поділяється на дві частини: покрівлю середнього мозку, розміщену над водопроводом середнього мозку (сільвієвим водопроводом), яка називається пластинкою покрівлі, або чотиригорбковою пластинкою, і ніжки мозку. Ніжки мозку заглиблюються у товщу середнього мозку; розділяються шаром сірої речовини, яка має значну кількість чорного пігменту меланіну і називається чорною речовиною, на дві частини — основу ніжок і покрив (мал. 21).