Полиненасыщенные жирные кислоты
Животные не способны синтезировать ни линолевую, ни линоленовую жирные кислоты из-за отсутствия соответствующих десатураз. Поэтому эти кислоты должны обязательно поступать с пищей. Полиненасыщенные жирные кислоты, например, арахидоновая, 20:4 (5,8,11,14), являются непосредственным предшественником простагландинов .
Синтез простагландинов |

Регуляция синтеза и окисления жирных кислот в печени
Переключение процессов синтеза жирных кислот на их окисление происходит при смене периода пищеварения на постабсорбтивное состояние и осуществляется с помощью регуляторных механизмов. Синтез малонил-СоА - ключевая реакция в регуляции синтеза и окисления жирных кислот . В период пищеварения в цитозоле увеличивается концентрация цитрата, который является переносчиком ацетильных остатков из митохондрий. Цитрат аллостерически активирует ацетил-СоА-карбоксилазу , что ускоряет синтез малонил-СоА и, следовательно, синтез жирных кислот. Малонил-СоА в свою очередь ингибирует ацил-карнитил-трансферазу , катализирующую перенос жирных кислот из цитозоля в митохондрии и "запускающую" механизм -окисления. Таким образом, увеличение концентрации малонил-СоА в период пищеварения "включает" процесс синтеза жирных кислот и "выключает" -окисление и синтез кетоновых тел. Ацетил-СоА-карбоксилаза также аллостерически ингибируется длинноцепочечными ацил-СоА, если они накапливаются, не успевая вступить в реакцию этерификации. Это пример ингибирования конечным продуктом процесса.
Аллостерическая регуляция метаболизма жирных кислот в печени |
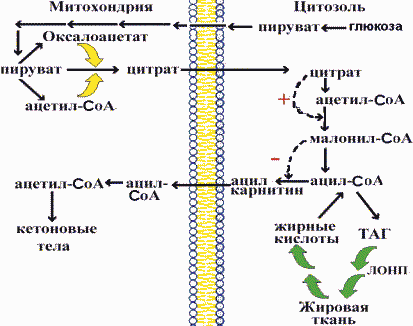
Кроме аллостерической регуляции существует гормональный контроль активности ацетил-СоА-карбоксилазы. Адреналин и глюкагон путем увеличения концентрации сАМР и активности протеинкиназы фосфорилируют ацетил-СоА-карбоксилазу и переводят ее в неактивное состояние. Эти гормоны также путем фосфорилирования переводят липазу в жировой ткани в активное состояние.
Гормональная регуляция обмена жирных кислот |

Следовательно, синтез жирных кислот прекращается, а начинается мобилизация ТАГ, окисление жирных кислот и синтез кетоновых тел, то есть включаются процессы, которые поставляют клеткам энергодативные вещества.
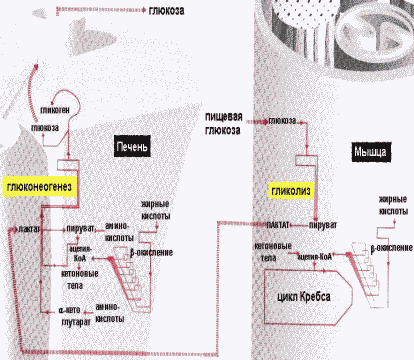
Взаимосвязь углеводного и жирового обмена |
Метаболизм холестерина и желчных кислот
Функции холестерина . Холестерин является предшественником в синтезе других стероидов: желчных кислот, стероидных гормонов, витамина D3. Холестерин входит как структурный компонент в состав мембран всех клеток. Существует два пути поступления холестерина:
из пищи животного происхождения (экзогенный холестерин)
синтез в печени (эндогенный холестерин)
Кроме печени в небольшом количестве холестерин может синтезироваться в клетках кишечника и кожи.
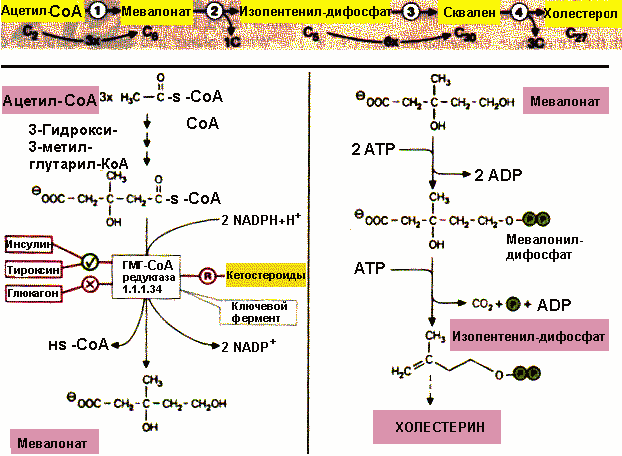
Биосинтез холестерина . Процесс происходит в цитозоле клетки. Молекула холестерина целиком "собирается" из ацетил-СоА. Промежуточным метаболитом является -окси- -метил-глутарил-СоА , а его восстановление в мевалоновую кислоту с использованием NADPH служит ключевой реакцией процесса. Скорость синтеза холестерина зависит от количества экзогенного холестерина, то есть поступающего с пищей. При поступлении 2-3 г холестерина в сутки синтез эндогенного холестерина подавляется.
Пути синтеза и использования холестерина |

Фермент гидроксиметелглутарил-СоА-редуктаза играет главную роль в регуляции синтеза холестерина. Холестерин подавляет синтез ГМГ-СоА-редуктазы и таким образом по механизму отрицательной обратной связи снижает скорость своего синтеза.
Синтез холестерина |