

Энергетика окисления жирных кислот
За один цикл -окисления образуется 1 молекула ацетил-СоА, окисление которого в цитратном цикле обеспечивает синтез 12 моль АТР. Кроме того, образуется 1 моль FADH2 и 1 моль NADH, при окислении которых в дыхательной цепи синтезируется соответственно 2 и 3 моль АТР (в сумме 5).
Таким образом, при окислении, например, пальмитиновой кислоты (С16) происходит 7 циклов -окисления, в результате которых образуется 8 моль ацетил-СоА, 7 моль FADH2 и 7 моль NADH+Н+. Следовательно, выход АТР составляет 35 АТР в результате -окисления и 96 АТР в результате цитратного цикла, что соответствует в сумме 131 моль АТР.
Образование кетоновых тел в печени (кетогенез)
-окисление жирных кислот в печени ведет к образованию восстановленных коферментов, которые используются для энергетических нужд клеток печени.
Основным путем дальнейшего использования ацетил-СоА, образованного при -окислении в печени, является синтез кетоновых тел , который происходит следующим образом: двухуглеродные молекулы конденсируются друг с другом с образованием в дальнейших реакциях ацетоацетата и -гидроксибутирата.
Синтез кетоновых тел |
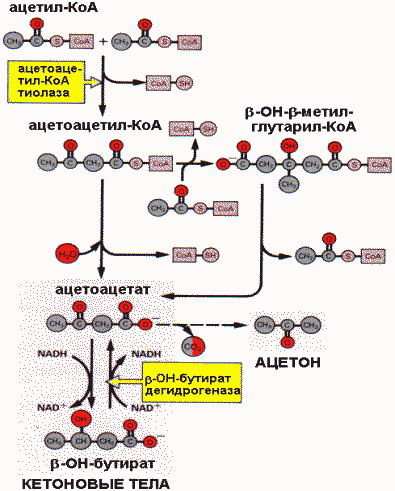
Эти две кислоты называются кетоновыми телами. Кетоновые тела диффундируют в кровь и используются внепеченочными тканями в качестве источников энергии. В норме концентрация кетоновых тел в крови 2 мг/дл. Ферменты, катализирующие синтез кетоновых тел, находятся в митохондриях. В определенных метаболических условиях, когда в печени происходит интенсивное окисление жирных кислот, образуется значительное количество кетоновых тел. Это происходит в состояниях, связанных с затруднениями в использовании углеводов, когда основным источником энергии для организма являются жирные кислоты (длительная мышечная работа, голодание, сахарный диабет). Скорость синтеза кетоновых тел может превышать потребление их тканями и вести к накоплению. Значительное увеличение концентрации кетоновых тел создает опасную ситуацию, которую называют кетоацидозом (кетозом) . При патологическом кетоацидозе ацетоацетат неферментативно декарбоксилируется, что сопровождается образованием ацетона, поэтому для кетоза характерен запах ацетона в выдыхаемом воздухе.
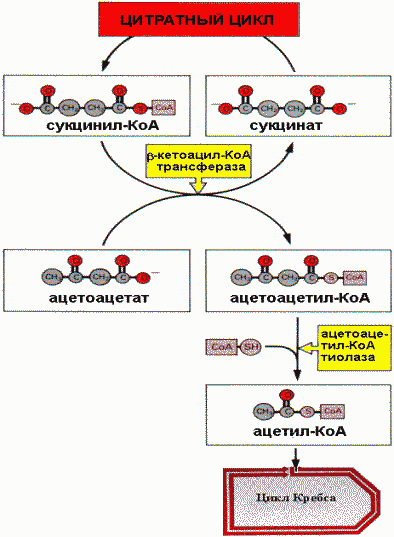
Окисление кетоновых тел |
Биосинтез жирных кислот
Процесс осуществляется в различных частях клетки. В цитоплазме синтезируются насыщенные жирные кислоты с углеродной цепью до С16 (пальмитат).
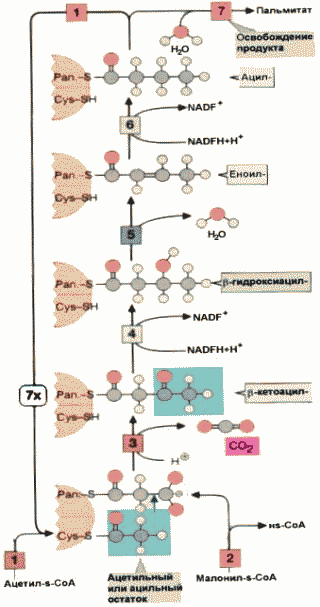
Синтез высших жирных кислот |
В митохондриях происходит дальнейшее наращивание цепи, а в ретикулуме насыщенные жирные кислоты превращаются в ненасыщенные, и также происходит удлинение цепи.
Основным продуктом системы синтеза, содержащимся в цитозоле, является пальмитиновая кислота . Субстратом для синтеза жирной кислоты служит ацетил-СоА , образующийся из глюкозы в результате окисления пирувата. Использование пирувата в синтезе пальмитата включает следующие метаболические пути: окисление глюкозы в процессе гликолиза до пирувата в цитозоле, затем окислительное декарбоксилирование пирувата в митохондриях и последующая конденсация ацетил-СоА с оксалоацетатом с образованием цитрата. Далее цитрат перемещается в цитозоль, где распадается на ацетил-СоА и оксалоацетат. Первая реакция в биосинтезе жирной кислоты - это карбоксилирование ацетил-СоА и превращение его в малонил-СоА. Реакция катализируется ацетил-СоА-карбоксилазой , коферментом которой является биотин . Затем следуют повторяющиеся циклы из шести реакций. Катализирует весь процесс пальмитилсинтетаза - полифункциональный белок - фермент, имеющий одну полипептидную цепь, упакованную в два домена . Оба домена имеют в своем составе активные центры, способные поочередно катализировать реакции каждого цикла. Ацильные группы связываются с одним из доменов, содержащим пантотеновую кислоту, а малонильные - с остатком цистеина другого домена. После инициации процесса путем реакции конденсации с образованием -кетоацил-фермента, растущая цепь жирной кислоты удерживается тиоэфирной связью с одним из доменов пальмитилсинтетазы. Ковалентно связанный субстрат поочередно попадает в активные центры фермента, где подвергается соответствующим превращениям. Очередной цикл синтеза начинается с присоединения новой молекулы малонил-СоА к одной из SH-групп активных центров доменов. Каждый цикл из шести реакций увеличивает длину цепи на два углеродных атома . Когда цепь достигает длины 16 углеродных атомов, тиоэфирная связь гидролизуется и пальмитат освобождается.
|
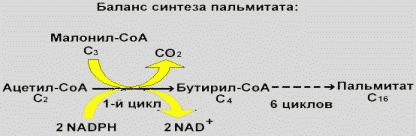
Суммарное уравнение синтеза пальмитата (7 циклов): Ацетил-СоА+7Малонил-КоА+14(NADPH+Н+) → пальмитат + 8HS-CoA+ 7CO2+14NADP++7H2O
Необходимый для восстановительных реакций NADPH образуется при окислении глюкозы по пентозофосфатному пути . Большинство жирных кислот имеют более длинную, чем пальмитат, углеводородную цепь, которая может содержать ненасыщенные связи. Следовательно, биосинтез большинства жирных кислот требует участия ферментов элонгации и десатурации . Активность этих ферментов связана с эндоплазматическим ретикулумом, но иногда может протекать и в митохондриях.