

Многообразие живых организмов – основа организации и устойчивости биосферы
Многообразие и разнообразие живых систем поражает воображение. За все время эволюции жизни на Земле существовало колоссальное количество различных видов живых организмов: 750 - 800 млн, - причем большая часть из них прекратила свое существование. В настоящее время различные источники дают разные цифры по количеству всех видов существующих ныне живых организмов: в одних фигурирует цифра в 2 млн, в других 3 - 5 млн. К концу прошлого тысячелетия насчитывалось около 1,2 млн видов животных и 0,5 млн видов растений. Большинство исследований и классификаций живых организмов свидетельствуют о том, что общее число видов растений и животных, обитающих в настоящее время на Земле, равняется примерно 5 млн, причем подавляющее большинство современных живых организмов - это тропические насекомые, многие из которых еще не изучены. Однако, по мнению Р. Мэя из Принстонского университета (Нью-Джерси, США), в приведенную схему расчетов следует внести коррективы, которые почти на порядок увеличат число организмов, существующих на Земле. Исследования обитателей тропических лесов показали, что только в тропиках имеется около 50 тыс. видов деревьев и примерно 30 млн видов насекомых. Речь пока не велась о простейших видах живых организмов на Земле, которых более 50 млн.
Лучше всего изучены живые организмы, проживающие в умеренной зоне Европы и Америки, причем в атмосфере, а хуже всего - живые организмы Арктики и Антарктики, а также глубинных слоев литосферы и гидросферы во всех зонах и на всех континентах. Наибольшее число древних видов, проживающих или произрастающих на Земле уже несколько десятков, а то и сотен тысячелетий, находится в Новой Зеландии.
Ежегодно исследователи описывают в среднем один новый вид млекопитающих и три вида птиц, причем в основном это обитатели тропиков, зоны, которая сейчас наиболее широко исследуется.
Совокупность всех живых организмов на Земле действует как единая мощная сила, преобразующая поверхностные слои нашей планеты. Кроме того, все единицы биосферы взаимодействуют между собой, все они одинаково необходимы для поддержания целостного и устойчивого состояния биосферы, роль которой в химических и энергетических процессах на Земле необыкновенно важна.
Биологическое разнообразие жизни на земле

Вирусы.
Вирусы – неклеточная форма жизни. Они являются облигатными паразитами, т.е. могут функционировать только внутри одно- или многоклеточного организма.
Первооткрыватель вирусов Д.И. Ивановский выявил два их основных свойства – они столь малы. Что проходят через фильтры, задерживающие бактерии, и их невозможно, в отличие от клеток, выращивать на искусственных питательных средах. Лишь с помощью электронного микроскопа удалось увидеть эти мельчайшие из живых существ ( от 20 до 3000 нм) и оценить многообразие их форм.
Вирусы – инфекционные агенты.
Своё название вирусы получили от латинского слова virus – яд. Ни один из известных вирусов не способен к самостоятельному существованию. Лишь попав в клетку, генетический материал вируса воспроизводится, переключая работу клеточных биохимических конвейеров на производство вирусных белков: как ферментов, необходимых для репликации вирусного генома – всей совокупности его генов, так и белков оболочки вируса. В клетке же происходит и сборка из нуклеиновых кислот и белков многочисленных потомков одного попавшего в неё вируса.
В зависимости от длительности пребывания вируса в клетке и характера изменения её функционирования различают три типа вирусной инфекции.
Если образующиеся вирусы одновременно покидают клетку, то она разрывается и гибнет. Вышедшие из неё вирусы поражают новые клетки. Так развивается литическая инфекция (греч. lysis – разрушение, растворение).
При вирусной инфекции другого типа, называемой персистентной (стойкой), новые вирусы покидают клетку хозяина постепенно. Клетка продолжает жить и делится, производя новые вирусы.
Третий тип инфекции называется латентным (скрытым). Генетический материал вируса встраивается в хромосомы клетки и при её делении воспроизводится и передается дочерним клеткам. При определённых условиях в некоторых из заражённых клеток латентный вирус активизируется, размножается, и его потомки покидают клетки. Инфекция развивается по литическому или персистентному типу.
Строение вирусов.
Вне зависимости от типа инфекции и характера заболевания все вирусы можно рассматривать как генетические элементы, одетые в защитную белковую оболочку и способные переходить из одной клетки в другую.
Отдельные вирусные частицы — вирионы представляют собой симметричные тела, состоящие из повторяющихся элементов. В сердцевине каждого вириона находится генетический материал, представленный молекулами ДНК или РНК. Велико разнообразие форм этих молекул: есть вирусы, содержащие двухцепочечную ДНК в кольцевой или линейной форме; вирусы с одноцепочечной кольцевой ДНК; одноцепочечной или двухцепочечной РНК; содержащие две идентичные одноцепочечные РНК.
Генетический материал вируса (геном) окружен капсидом — белковой оболочкой, защищающей его как от действия нуклеаз — ферментов, разрушающих нуклеиновые кислоты, так и от воздействия ультрафиолетового излучения. Капсиды состоят из многократно повторенных полипептидных цепей одного или нескольких типов белков. В основе взаимодействия вирусных белков друг с другом и с нуклеиновой кислотой лежит закон термодинамики, гласящий, что устойчивость системы приобретается при достижении минимального уровня свободной энергии. Для каждого вируса существует свой набор белков, который при сборке вириона дает оптимальную в энергетическом плане форму капсида. Большинство вирусов построены по одному из двух типов симметрии — спиральной или кубической.
Вирионы со спиральной симметрией имеют форму продолговатых палочек. В центре находится спирально закрученная нуклеиновая кислота (рис. 2). Капсид состоит из идентичных субъединиц белка, спирально расположенных вдоль молекулы нуклеиновой кислоты. По спиральному типу симметрии построено большинство вирусов, поражающих растения, и некоторые вирусы бактерий, так называемые бактериофаги или просто фаги.
Большая часть вирусов, вызывающих инфекции у человека и животных, имеет кубический тип симметрии. Капсид почти всегда имеет форму икосаэдра – правильного двадцатигранника с двенадцатью вершинами из равносторонних треугольников (рис. 3).
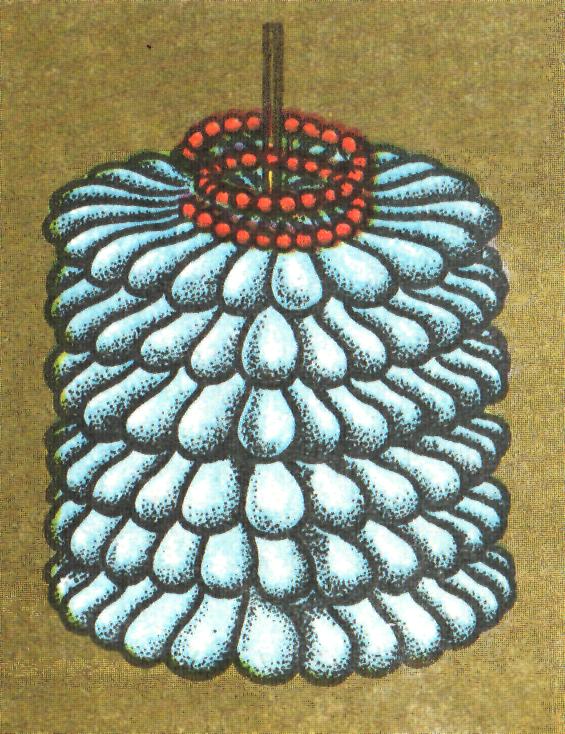
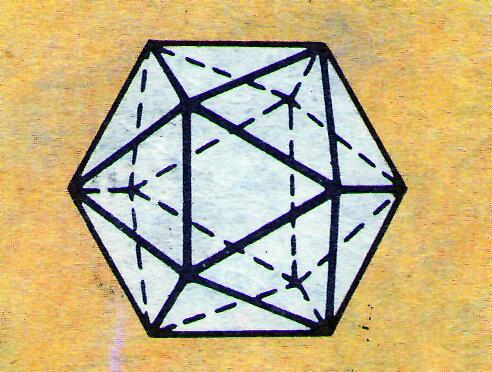
Рис. 2. Модель части ВТМ Рис. 3. Модель икосаэдра
Существуют вирусы и с более сложным строением. Некоторые фаги помимо икосаэдрической головки, содержащей генетический материал, имеют полый цилиндрический отросток, окруженный чехлом из сократительных белков и заканчивающийся шестиугольной площадкой с шестью короткими выростами и шестью длинными фибриллами — нитями. Такая сложная конструкция обеспечивает впрыскивание генетического материала фага внутрь бактериальной клетки.
Многие вирусы помимо белкового капсида имеют внешнюю оболочку. Кроме вирусных белков и гликопротеинов (белков, ковалентно связанных с углеводными молекулами) она содержит еще и липиды, позаимствованные из плазматической мембраны клетки-хозяина. Вирус гриппа - пример спирального вириона в оболочке с кубическим типом симметрии.
Современная классификация вирусов основана на виде и форме их нуклеиновой кислоты, типе симметрии и наличии или отсутствии внешней оболочки.
Размножение вирусов. Размножение вирусов включает в себя три процесса: репликацию вирусной нуклеиновой кислоты, синтез вирусных белков и сборку вирионов.
Разнообразие видов и форм вирусных нуклеиновых кислот определяет и разнообразие способов их репликации. Бактериофаг Т4 имеет одну двухцепочечную линейную молекулу ДНК, состоящую из 160·103 пар нуклеотидов. В ней закодировано более 150 различных белков, в том числе более 30 белков, участвующих в репликации фаговой ДНК. Обезьяний вирус SV40 имеет двухцепочечную кольцевую ДНК. У вируса оспы две комплементарные цепи линейной ДНК на обоих концах соединены одна с другой ковалентной фосфодиэфирной связью. Этот самый крупный из известных вирусов содержит более 240 генов. Репликация у вирусов с двухцепочечной ДНК принципиально не отличается от репликации бактериальной или эукариотической ДНК.
Паразитирующие в бактериальных клетках Е. coli фаги М13 и λХ174 имеют кольцевую одноцепочечную ДНК. В зараженной клетке бактериальные ферменты репликации синтезируют комплементарную ей цепь, которая служит матрицей для образования фаговых ДНК. Они соединяются с фаговыми белками, также синтезированными бактериальными ферментами, и новые фаги покидают клетку-хозяина. Многие вирусы растений содержат одну линейную молекулу РНК, например первый из описанных вирус табачной мозаики (ВТМ). Молекула РНК ВТМ заключена в белковый капсид, состоящий из 2130 идентичных полипептидных субъединиц. В 1955 г. в изящном эксперименте с «переодеванием» Френкель-Конрад впервые показал, что РНК может выполнять функцию носителя генетической информации. (Для ДНК эта функция была доказана раньше.) Он взял два штамма ВТМ, дающих различные картины поражения листьев табака. Отделив белки от РНК, он реконструировал вирионы таким образом, чтобы РНК из одного штамма покрывалась белковым чехлом другого. Картина поражения листьев, зараженных реконструированным вирусом, не зависела от того, какому штамму принадлежали белки, она определялась лишь РНК.
Репликация РНК вируса табачной мозаики осуществляется ферментом, называемым РНК-зависимой РНК-полимеразой, закодированной в геноме вируса. Сначала этот фермент строит комплементарную цепь РНК, так называемую минус-цепь (она не кодирует белки в отличие от вирусной РНК, кодирующей белки и поэтому называемой плюс-цепью), а затем по ней, как по матрице, синтезирует множество вирусных РНК.
У так называемых вирусов с «негативными» геномами, к которым относится вирус гриппа, инфицирующая РНК является минус-цепью и не кодирует белков. Только комплементарная ей плюс-цепь РНК, синтезирующаяся в зараженных вирусом клетках, несет информацию о создании новых вирусных частиц. Она же служит матрицей для образования большого количества вирусных минус-цепей РНК.
Две одинаковые одноцепочечные молекулы РНК внутри икосаэдрического белкового капсида содержат некоторые онкогенные (опухолеродные) вирусы. Они имеют еще и внешнюю оболочку, состоящую из двойного липидного слоя плазматической мембраны клетки-хозяина, а также белков и гликопротеинов вирусного происхождения. Такое же строение имеет вирус иммунодефицита человека (ВИЧ), вызывающий синдром приобретенного иммунного дефицита (СПИД).
Из онкогенных вирусов первым был открыт вирус саркомы Рауса (ВСР), вызывающий злокачественные опухоли у кур. Изучение механизма трансформации клетки, т. е. превращение из нормальной в раковую, привело в 1970 г. американских ученых Г. Темина и Д. Балтимора к открытию явления обратной транскрипции. ВСР содержит фермент, называемый обратной транскриптазой. Он представляет собой ДНК-полимеразу, которая сначала синтезирует цепь ДНК, используя в качестве матрицы одну из идентичных молекул вирусной РНК, а зачтем вторую, комплементарную цепь ДНК. В результате образуется двухцепочечная ДНК. Она может встроиться в хромосому клетки-хозяина. Такой процесс встраивания в хромосомную ДНК называют интеграцией. Вирусный геном в форме интегрированной ДНК, синтезированной по проникшей в клетку вирусной РНК с помощью обратной транскриптазы, называется провирусом.
Провирус становится частью генетического материала клетки, реплицируется вместе с клеткой ДНК и при делении передется дочерним клеткам. В скрытой (латентной) форме провирус может пребывать бесконечно долгое время, переходя от родителей к потомкам через сперматозоид или яйцеклетку.
Канцерогенные, т.е. приводящие к раку, факторы, такие, как рентгеновые лучи, табачный дым, асбестовая пыль, некоторые продукты переработки нефти, бензол и другие, могут активировать провирус в отдельных клетках. В них образуются вирусные РНК и белки, происходит злокачественная трансформация. Раковые клетки отличаются от нормальных тремя признаками: 1) они быстрее делятся, затрачивая большое количество энергии молекул АТФ; 2) они частично дедифференцируются, т.е.утрачивают часть признаков, приобретённых клетками в процессе развития организма, и становятся похожими на зародышевые клетки; 3) они иногда теряют присущую в норме способность к тесному сцеплению с соседними клетками, а поэтому могут отделятся от них, перемещаться в другие части тела и давать начало новым опухолям, т.е. метастезировать.
РНК-содержащие вирусы, являющиеся первопричиной злокачественной трансформации, называют онкогенными ретровирусами (лат. retro – возврат назад) из-за того что обратная транскрипция – необходимый этап в их размножении.
Вирусы — факторы изменения генетической информации организмов. При нормальной транскрипции вирусной ДНК, интегрированной в хромосому клетки, могут транскрибироваться и расположенные рядом хозяйские гены. При обратной транскрипции вирусной РНК эти гены могут встроиться в хромосомы другого организма и оказаться в необычном для них окружении. Тем самым ретровирусы могут не только переносить гены, привнося в организмы дополнительную наследственную информацию, но и изменять работу хозяйских генов. Ретровирусы могут переносить гены между клетками одного организма, между организмами не только одного, но и разных видов или классов, когда половая гибридизация исключена. Поток генов между далекими организмами представляется очень реальным - на это указывает сходство у самых разных животных генов, одновременно входящих в состав ретровирусов. Сегодня вирусы рассматривают не только как возбудителей инфекционных болезней, но и как переносчиков генетической информации между видами. Кроме того, сам факт попадания вируса в живую клетку и перестраивание ее биохимических конвейеров на создание вирусного потомства если не убивает клетку, то не проходит для нее бесследно. Разрывы хромосом, изменения в порядке расположения генов, а также изменения в самих генах остаются в «генетической памяти» клеток, посещенных незваными пришельцами.