Российский национальный исследовательский медицинский университет имени Н. И. Пирогова
Кафедра Биологической химии
Реферат на тему:
Вторичные посредники. Типы и роль во внутриклеточной передаче сигнала
Выполнил студент 2-го курса 213 группы Лечебного факультета Парсаданян Гайк Каренович
2012г.
г. Москва
Оглавление
Введение………………………………………………………………………………………………………………..….. 2
Мембранные рецепторы………………………………………………………………………………………….. 4
Виды мембраносвязанных рецепторов………………………………………………… 5
Структура мембранных рецепторов………………………………………………………. 6
Основные этапы передачи гормональных сигналов……………………………………………... 6
ГТФ-связывающие белки (G-белки)…………………………………………………………………..…….. 7
Вторичные посредники и механизмы их действия………………………………………….……. 8
Циклический АМФ……………………………………………………………………………..…………... 9
G-белок и аденилатциклаза…………………………………………………………..………. 9
Синтез и расщепление цАМФ………………………………………………………….……. 11
Этапы передачи сигнала………………………………………………………………….……. 12
цАМФ-зависимая протеинкиназа…………………………………………………….…… 14
цАМФ-регулируемые протеинфосфатазы…………………………………………….. 14
Биохимическая модель действия цАМФ……………………………………………… 15
Множественный эффект цАМФ…………………………………………………………….. 16
Са2+ - вторичный посредник…………………………………………………………………….……. 17
Белки, связывающие кальций………………………………………………………..….…. 18
Механизм действия……………………………………………………………………………….. 19
Процессы или ферменты, регулируемые комплексом Ca2+ с кальмодулином……………………………………………………………….………. 20
Взаимодействие цАМФ и Са2+………………………………………………………………. 21
Фосфоинозитидный путь передачи сигнала……………………………………………….. 22
Механизм действия……………………………………………………………………………….. 22
Фосфолипаза С………………………………………………………………………………………… 24
Инозитолтрифосфат………………………………………………………………………………… 25
Диацилглицерол…………………………………………………………………………………….. 25
Протеинкиназа С…………………………………………………………………………………….. 25
Множественные эффекты фосфоинозитидного пути…………………………… 26
Циклический ГМФ…………………………………………………………………………………………… 26
Заключение……………………………………………………………………………………………………….………. 28
Список литературы……………………………………………………………………………………………….…… 30
Введение
Одним из важнейших этапов прогрессивной эволюции явилось возникновение многоклеточных организмов, у которых разные типы тканей выполняют различные функции. Клетки любого многоклеточного организма нуждаются в обмене информацией друг с другом – для регуляции своего развития и организации в ткани, для контроля процессов роста и деления и для координации функций – одним словом, для выживания организма в целом. У многоклеточных животных сформировались два вида такой связи. Один вид – быстрая передача электрохимических сигналов нервной системой из одной части организма в другую. Второй вид связи основан на том, что нервная система и другие специализированные ткани выделяют в общую циркуляцию особые вещества для передачи сигналов отдаленным органам. Без таких молекулярных посредников и регуляторов была бы невозможна ежеминутная и повседневная координация функций различных органов и тканей. Подобно тому, как мышечная активность невозможна без координации и контроля, осуществляемых нервной системой с помощью электрических сигналов, так и рост, поддержание стационарного состояния и размножение невозможны без координации и контроля, осуществляемых внеклеточными и внутриклеточными посредниками.
Можно сказать еще, что взаимодействие животных клеток осуществляется следующими тремя способами:
Клетки выделяют химические вещества, служащие сигналами для других клеток, расположенных на некотором расстоянии.
Клетки несут на своей поверхности связанные с плазматической мембраной сигнальные молекулы, оказывающие влияние на другие клетки при непосредственном физическом контакте.
Клетки образуют щелевые контакты, прямо соединяющие цитоплазму двух взаимодействующих клеток, что делает возможным обмен малыми молекулами.
В любом случае, нас интересует тот путь коммуникации на расстоянии, которая осуществляется с помощью секретируемых химических сигналов. И тут можно добавить, что химические сигнальные механизмы различаются по расстояниям, на которых они действуют:
Аутокринная регуляция - клетка выделяет вещества, которые оказывают специфическое действие на ту же клетку.
Паракринная регуляция – клетки выделяют локальные химические медиаторы, которые поглощаются, разрушаются или иммобилизируются так быстро, что успевают подействовать путем диффузии только на клетки ближайшего окружения.
Нейрокринная регуляция – выделение нейронами нейромедиаторов в специализированные межклеточные контакты, называемые химическими синапсами; данный способ сигнализации используется только в нервной системе.
Эндокринная сигнализация – специализированные эндокринные клетки выделяют гормоны, которые разносятся кровью и воздействуют на клетки-мишени, находящиеся иногда в самых разных частях организма; если при этом посредник секретируется в кровь нервными клетками, то речь идет о функции нейроэндокринной системы.

В каждом случае мишень реагирует на определенный внеклеточный сигнал с помощью специальных белков, называемых рецепторами, которые связывают сигнальную молекулу и инициируют ответ. Главные различия между этими путями касаются быстроты и избирательности воздействия сигнала на определенные мишени. К примеру, нейромедиаторы – посредники, выделяемые нервными клетками, – действуют на небольшом расстоянии, активируя рецепторы постсинаптических клеток. Гормоны же передают сигнал на более далекие расстояния, обычно через кровеносную систему и межклеточную жидкость, поэтому сигнал передается гораздо медленнее.
Мембранные рецепторы
Для регуляции деятельности клетки с помощью гормонов, находящихся в плазме крови, необходимо обеспечить возможность клетки воспринимать и обрабатывать этот сигнал. Эта задача усложняется тем, что сигнальные молекулы (нейромедиаторы, гормоны) имеют разную химическую природу, реакция клеток на сигналы должна быть различной по направленности и адекватной по величине.
В связи с этим, эволюционно сформировались два основных механизма действия сигнальных молекул по локализации рецептора:
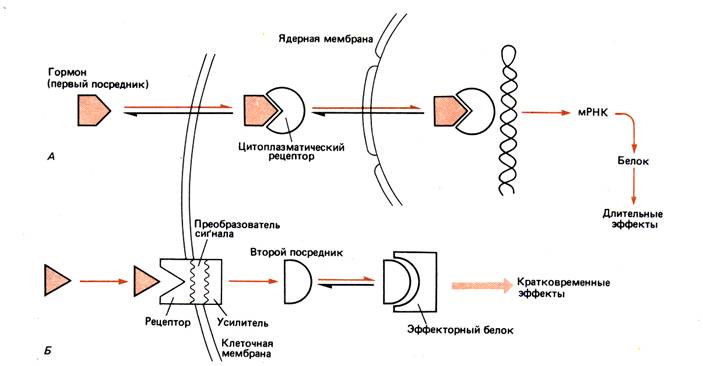
Цитозольный – рецептор расположен в цитозоле. По данному механизму работают стероидные и другие липофильные гормоны; по–видимому, воздействуют внутри клетки на генетический аппарат. Эти гормоны называют внеклеточными и первыми посредниками.
Мембранный – рецептор расположен на мембране. Для этих рецепторов в зависимости от способа передачи гормонального сигнала в клетку выделяют три вида мембраносвязанных рецепторов и, соответственно, три механизма передачи сигнала. По данному механизму работают пептидные и белковые гормоны, катехоламины, эйкозаноиды. Проявляются более быстрые и кратковременные реакции, влияя, как правило, на обмен веществ. Действие гормонов прекращается вскоре после их недолговременного присоединения к белковым рецепторным молекулам на поверхностных мембранах клеток–мишеней. Взаимодействие гормона с рецептором ведет к запуску внутриклеточного каскадного механизма (т. е. к цепи реакций, усиливающих исходный сигнал), что приводит к образованию внутриклеточного регулятора в цитозоле клетки. Молекулу (или комплекс молекул) внутриклеточного регулятора, образующуюся после взаимодействия гормона с рецептором, называют внутриклеточным или вторым посредником.
На следующем рисунке:
Жирорастворимый гормон проходит через мембрану и, присоединившись внутри клетки к молекуле рецептора, образует активный комплекс, который воздействует на генетический аппарат.
Гормон, нерастворимый в жирах, присоединяется к рецептору на поверхности клетки и стимулирует ферментативное образование второго посредника, который, связавшись с другой молекулой, образует с ней метаболически активный комплекс.
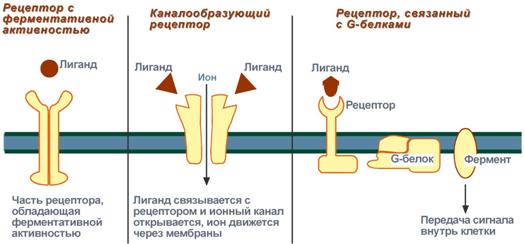
Виды мембраносвязанных рецепторов
Рецепторы, обладающие каталитической активностью – при взаимодействии лиганда с рецептором активируется внутриклеточная часть (домен) рецептора, имеющий тирозинкиназную или тирозинфосфатазную или гуанилатциклазную активность. По этому механизму действуют инсулин, пролактин, интерлейкины, ростовые факторы, интерфероны α, β, γ.