

93. Механизм фиксации азота бобовыми, стадии и значение нодуляции.
N2 – самый обильный газ атмосферы (70%), но он не доступен растениям.
При фиксации азота N2 переводится в NH3
Симбиотические взаимоотношения растений с азотфиксирующими бактериями позволяют растениям получить «встроенный» источник азота, т.е. Производить фиксацию азота в своем «организме».
Наиболее важные симбиозы с азотфиксирующими бактериями образуют бобовые (горох, бобы и др.).
Корни бобовых разбухают, образуя так-называемые клубеньки или «нодулы», состоящие из клеток растения и N2-фиксирующих бактерий рода Rhizobium
Внутри корневых клубеньков Rhizobium живут в специальных везикулах и называются бактероидами.
Rhizobium – грам-отрицательные, подвижные палочки.
Бактерии получают из корня сахара и все необходимое для жизнедеятельности, растение получает фиксированный азот в виде аммиака
(NH3 / NH4+).
NH4+ транспортируется через неселективные катионные каналы перибактероидной мембраны (она окружает бактероид) в клетку растения.
Нитрат, попадая в корневые клетки, восстанавливается до аммония, который затем включается в аминокислоты.
Аммоний, «синтезированный» из нитрата клетками быстро переходит в аминокислоты, которые даже в больших количествах не токсичны для растения.
В клетках существует совершенная система метаболической регуляции, набор согласованно работающих ферментов, обеспечивающих утилизацию азота.
Одни из них также не позволяют накапливаться избыточному аммонию, ограничивая восстановление нитратов.
Если же по каким либо причинам он всё же появляется, другие ферменты связывают его в форме обогащённых азотом запасных соединений (высокоазотные аминокислоты, амиды или мочевина).
94. Химизм и физиологическая значимость реакций переаминирования.
Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937 г. биохимиками А. Е. Браунштейном и М. Г. Крицман.
Реакция заключается в межмолекулярном переносе аминогруппы с аминокислоты на кетокислоту и катализируется ферментами, называемыми аминотрансферазами.
Наиболее интенсивно протекают следующие реакции переаминирования:
1. глютаминовая кислота + щавелевоуксусная кислота
аминотрансфераза
L – кетоглютаровая кислота + аспарагиновая кислота
2. глютаминовая кислота + пировиноградная кислота
аминотрансфераза
L – кетоглютаровая кислота + L – аланин
3. аспарагиновая кислота + пировиноградная кислота
аминотрансфераза
щавелевоуксусная кислота + L – аланин
Реакция переаминирования идет в несколько этапов:
1. Образование комплекса между реагирующей аминокислотой и фосфопиридоксалем аминотрансферазы.
2. Внутримолекулярная перегруппировка образовавшегося комплекса, распадающегося далее на соответствующую аминокислоте кетокислоту и фосфопиридоксаминовую форму аминотрансферазы, в которой кофермент представляет собой фосфопиридоксамин.
3. Фосфопиридоксаминовая форма аминотрансферазы затем реагирует с участвующей в переаминировании кетокислотой, образуя новый комплекс.
4. Образовавшееся комплексное соединение также подвергается внутримолекулярной перегруппировке, после чего оно распадается на новую аминокислоту и исходную пиридоксалевую форму аминотрансферазы.
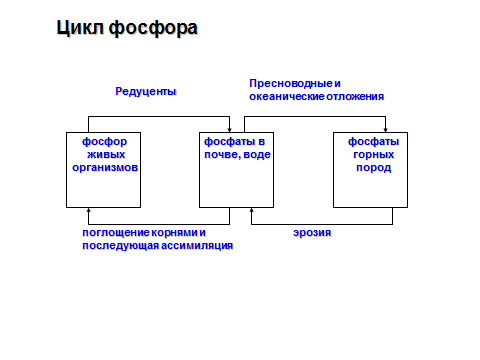
95.Роль фосфора и серы в жизни растений. Циклы фосфора и серы.
Фосфор: Синтез нуклеиновых кислот, АТФ и некоторых белков. Входит в состав фосфолипидов мембран, играет ключевую роль в переносе энергии
Он входит в состав протоплазмы и клеточного ядра. Фосфор играет большую роль в иммунитете растений и сохранении материнских признаков у потомства, так же накапливается в семенах и служит резервом при его прорастании. Он играет большую роль в фотосинтезе, способствует равномерному плодоношению и ускоренному росту.
Общие признаки фосфорного голодания у растений такие: лист приобретает фиолетовый или пурпурный цвет, на краях нижних листьев появляются желто-бурые или бурые пятна, они иногда закручиваются. Цветение и созревание задерживается, ухудшается рост,
молодые листья мелкие, отходят от веток под острым углом.
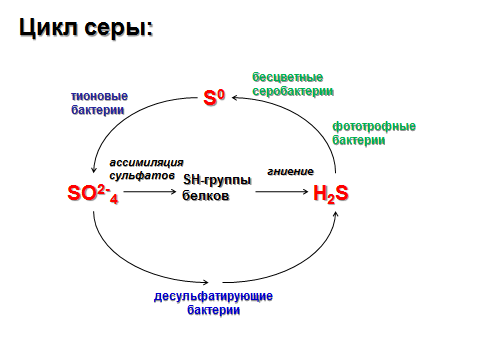
Сера: Синтез белков и многих органических соединений, например кофермента А, принимает участие в ЭТЦ дыхания и фотосинтеза.
Функции серы:
1.входит в состав белков, аминокислот, растительных масел, витаминов, антибиотиков.
2.играет большое значение в окислительно-восстановительных процессах, проходящих в растениях.
3.способствует фиксации азота из атмосферы.
Источник питания Серой – соли серной кислоты. Чся листьями из воздуха в виде SO2
В молодых растущих органах растений сера находится в основном в
восстановленной форме. По мере старения растений возрастает количество окисленной формы соединений Серы.При недостатке задерживается синтез белков. Признаки недостаточности Серы сходны с признаками азотного голодания. Развитие растений замедляется, уменьшается размер листьев, удлиняются стебли, листья и черешки становятся деревянистыми.При серном голодании листья не отмирают, хотя окраска их становится бледной.Потребность в Сере у разных растений неодинакова. Больше всего её содержится в бобовых, подсолнечнике, горчице, капусте.
Основными особенностями круговорота фосфора, таким образом, являются:
отсутствие атмосферного переноса;
наличие единственного источника – литосферы;
тенденция к накоплению в конечных водоёмах стока.