

Ооморфологическая характеристика различных кладок дрофы в северной части н. Поволжья (сост. По [488, 489, 491])31
Признак |
n |
M m |
CV, |
Lim |
t-критерий Стьюдента |
L (длина), мм |
24 |
79.8 0.60 |
4.70 |
71.6–86.0 |
0.964/0.391 |
74 |
79.1 0.41 |
5.65 |
73.1–92.4 |
0.964/0.634 |
|
39 |
79.5 0.48 |
5.23 |
71.8–87.0 |
0.391/0.634 |
|
B (максимальный диаметр), мм |
24 |
55.7 0.30 |
3.24 |
53.0–58.5 |
1.515/0.993 |
74 |
56.3 0.26 |
3.48 |
49.3–61.8 |
1.515/0.535 |
|
39 |
56.1 0.27 |
1.74 |
53.8–58.7 |
0.993/0.535 |
|
M (масса), г |
35 |
133.3 1.12 |
4.22 |
117.2–141.1 |
- |
25 |
136.3 1.39 |
2.28 |
118.6–148.0 |
1.681 |
|
Sph (индекс округлости), |
24 |
70.3 0.58 |
4.82 |
63.2–75.8 |
1.418/1.154 |
74 |
71.3 0.46 |
6.84 |
58.9–80.0 |
1.418/0.419 |
|
39 |
71.1 0.38 |
4.38 |
65.1–76.3 |
1.154/0.419 |
|
V (обьем яйца), мл |
24 |
128.4 2.42 |
8.99 |
107.7–142.6 |
-/0.071 |
74 |
128.4 1.37 |
8.03 |
97.0–156.6 |
-/0.101 |
|
39 |
128.6 1.42 |
7.70 |
110.9–142.1 |
0.071/0.101 |
|
Iw (индекс массы) |
35 |
2.99 0.005 |
2.20 |
2.84–3.03 |
- |
25 |
3.03 0.008 |
1.21 |
2.99–3.17 |
4.255 |
Примечание. Кладка, состоящая из одного (*),двух (**) и трех (***) яиц.
В отношении других ооморфологических показателей нами и другими авторами на протяжении нескольких десятилетий приводились несколько различающиеся значения, что обусловлено постепенным увеличением объема анализируемой выборки. В конечном итоге на основе анализа максимально возможного количества данных были получены сведения, в которых между показателями индекса массы и формой отдельных яиц дрофы была выявлена статистически достоверная связь. Так, чем больше размеры яйца, его масса, тем в среднем больше его коэффициент массы (r = 0.626 при p 0.05). Еще более сильная корреляция отмечена между индексом округлости яйца и коэффициентом его массы: чем «круглее» яйцо, тем в среднем больше его коэффициент массы (r = 0.776 при p 0.01). В то же время анализ оологических данных с использованием коэффициента массы (0.03 0.001) показал, что в начале гнездования отклоняющиеся по форме и массе яйца составляют не более 3–4.5% от общего количества отложенных яиц, а в составе поздних кладок таких яиц более 12% [457]. Объем яиц из саратовской популяции составляет 91.8–149.5, в среднем 128.6 ± 1.75 мл [470], их масса – 115.4–146.9, в среднем 134.8 ± 1.65 г [490].
Таким образом, анализ полученных многолетних данных показал, что в условиях северной части Н. Поволжья в среднем величина кладки является постоянной. При этом наиболее крупные и массивные яйца с эллипсоидной, укороченно- или удлиненно-эллипсоидной формой характерны для кладок, состоящих из трех яиц. По-видимому, они откладываются самками в оптимальном репродуктивном возрасте.
Яйца откладываются с интервалом в 26–50 ч, насиживание начинается с первого яйца. Насиживает кладку исключительно самка. В период непрерывного ночного насиживания кладки самка периодически меняет позу, чистит свое оперение или зондирует клювом дно гнезда. В светлое время суток вне насиживания интервалы между различными составляющими ее поведения во время кормовой паузы составляют в среднем 17.4 мин (lim 4–32). Кроме того, продолжительность «активного периода суток» увеличивается при повышении температуры воздуха. При значительных температурах начало активности приходится на более ранние утренние часы и, наоборот, при низкой температуре окончание непрерывного ночного насиживания отодвигалось на более поздний срок. Общая продолжительность регулярного насиживания – 24–28 дней, но при частом беспокойстве птиц она может увеличиться до 31 дня [23].
Кормится самка 3–4 раза в сутки в наиболее теплые часы. Птица в это время очень осторожна, при малейшей опасности возвращается к гнезду и прикрывает собой кладку. На приближающих людей, хищных млекопитающих, домашних животных реагируют за 100–300 м. Насиживающая самка первоначально затаивается, но если «опасность» продолжает приближаться к гнезду, покидает его и, стараясь быть незамеченной, отходит на 30 м и более, после чего взлетает.
К настоящему времени достаточно подробно изучены вопросы динамики основных оологических показателей в ходе искусственной инкубации [498]. Например, на основе морфологического исследования 35 яиц, собранных из гибнущих при сельскохозяйственных работах кладок дрофы, было установлено, что изменения индексов массы яиц (Iw) линейно зависят от дня инкубации (С) и описываются следующими уравнениями: Iw = 3.308 – 0.024 × С (оплодотворенные) и Im = 3.308 – 0.0401 × С (неоплодотворенные). Движение яиц начинает фиксироваться в основном на 25-е сутки (33.3%), писк птенцов в яйце – на 27-е (51.7%), наклев дрофенок делает на 27-е (51.8%) сутки [490].
От появления трещин до вылупления птенца проходит от 16 до 24 ч; появляются птенцы с интервалом в среднем в 24 ч [400, 409]. В условиях искусственной инкубации в 2001 г. интервал между вылуплением птенцов в кладках, состоящих из двух яиц, в среднем составил 33 ч; в кладках из трех яиц между вылуплением первого и второго птенца – 24 ч, второго и третьего – 20 ч. Бюджет времени в процессе вылупления птенцов распределяется следующим образом: промежуток между движением и писком – 57 ч, писком и наклевом – 12 ч, наклевом и вылуплением – 24 ч. Хорошую выводимость имеют яйца, взятые на искусственную инкубацию как на поздней, так и на ранней стадиях насиживания [490].
Не принимающая участия в размножении часть популяции концентрируется на богатых в трофическом отношении участках, где формируется стайная структура. В данной ситуации возможно образование однополых групп из самок или самцов, состав которых на протяжении всего периода насиживания постоянен. Встречи групп самок, не размножающихся в данном полевом сезоне или потерявших кладки (до 52%), регистрируются с последней декады мая – первых чисел июня. Обычно они держатся на изучаемой территории на протяжении всего репродуктивного периода. Например, данные спутникового слежения свидетельствуют о том, что размеры участка локализации самки, потерявшей кладку, составляли в период с мая по октябрь включительно 5.3 × 6.6 км с центром в месте отлова [27]. К середине июля из-за низкого успеха размножения число самок в однополых группах постепенно нарастает и составляет на данный период около 68%. В этот же период наблюдается перемещение дроф на поля яровых, посевы проса, суданской травы и поля люцерны посевной (Medicago sativa); озимые посевы не используются птицами в качестве кормовых местообитаний [26].
Первые птенцы в условиях саратовского Заволжья наблюдаются с середины мая, а массовое появление выводков регистрируется в начале июня [26]. После вылупления самка разбивает оставшиеся половинки скорлупы клювом и съедает их. После появления первого птенца она часто оставляет гнездо и перемещается на короткие расстояния с птенцом, которого кормит с клюва нежными частями растений. Выводок покидает гнездо через несколько часов после вылупления второго (последнего) птенца и уже в первый день может отходить от него на сотни метров [23]. Однако существует мнение, что самка держится с птенцами на участке гнездования недалеко от гнезда достаточно долгое время. Например, О.С. Опарина и М.Л. Опарин [26] наблюдали самку с двумя птенцами 10–15-дневного возраста на одном месте 2 ночи подряд. Выводки дроф держатся исключительно скрытно, избегая, однако, мест с густым высоким травостоем (в частности, озимую рожь – Secale cereale или посевную люцерну), где птенцам трудно передвигаться и где всегда относительно сыро и жарко. Излюбленными местами пребывания птенцов в условиях Саратовской области являются посевы ячменя, проса, житняка (и особенно зарастающие пары (если задержана их обработка).
С первых часов жизни птенцы чутко реагируют на звуковые сигналы родителей, а через 5–8 ч после вылупления у них появляются хватательные движения, после чего самка начинает их кормить. Но в целом первое время они малоподвижны, долго лежат, лишь появление самки с кормом вызывает активную реакцию. Как показали наши наблюдения, в течение первого дня она кормит их остатками скорлупы, подбирая кусочки размерами не более 3–4 мм и передавая их птенцам с клюва. С 2–5-го дня птенцы двигаются больше, пытаются самостоятельно брать корм, реагируют на пролетающих близко насекомых, в случае опасности затаиваются. Однако самка продолжает кормить их в течение 3 недель, после чего у них происходит замена хватательных движений склевывающими [400, 408].
В первые несколько недель после вылупления суточная активность взрослых дроф и птенцов характеризуется наличием двух пиков – утреннего (с рассвета до 10 ч) и вечернего (с 18 ч и до сумерек). В пасмурные дни четких пиков активности не выявлено. Однако экстремальные погодные условия (резкое понижение температуры, сильный дождь, ветер) временно изменяют ритм суточной активности и продолжительность отдыха. Как правило, в таких случаях молодые и взрослые птицы активизировались позже обычного времени, сокращая таким образом «рабочий день». В возрасте пяти недель птенцы поднимаются на крыло, однако полной самостоятельности достигают лишь в период первой зимовки [389, 400]. Масса только что вылупившегося дрофенка составляет от 83.2 до 103.7 г (в среднем 94.4 г), что составляет 82.8% массы насиженного яйца [470]. Наиболее интенсивно масса тела птенцов начинает увеличиваться с 5–10-дневного возраста (рис. 20).
Д
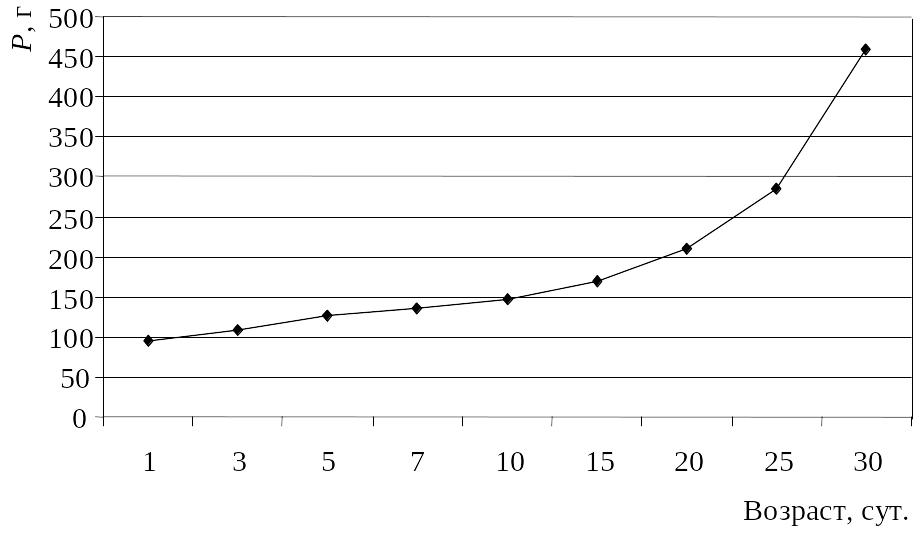 Рис.
20. Динамика
увеличения массы птенцов дрофы в
условиях искусственного выращивания
Рис.
20. Динамика
увеличения массы птенцов дрофы в
условиях искусственного выращивания
О
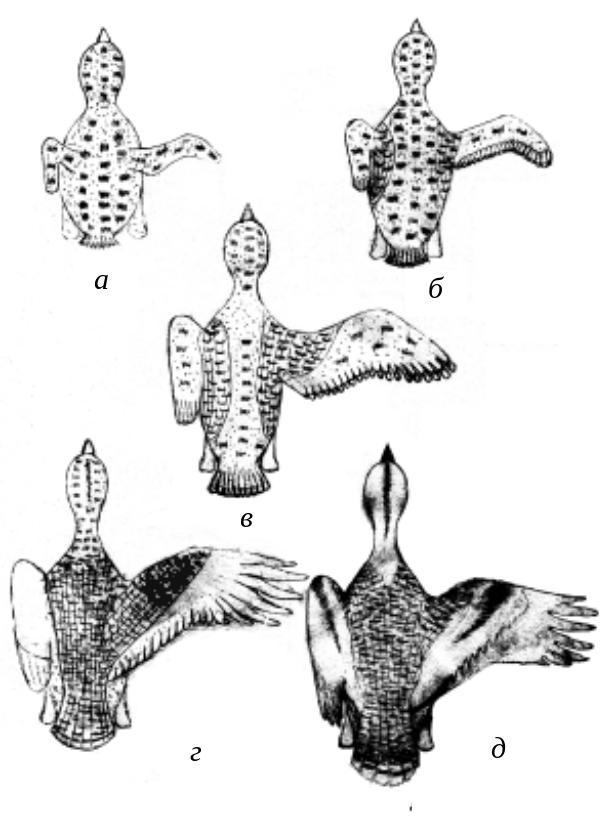 Рис.
21. Схема развития оперения у птенцов
дрофы
Рис.
21. Схема развития оперения у птенцов
дрофы
В сентябре – октябре, т.е. в возрасте более 4 месяцев, начинается полная линька, определяющая появление первого промежуточного наряда [389]. При этом сменяется оперение на голове и теле, кроющие крыла, часть внутренних первостепенных и третьестепенных маховых, центральные рулевые, а остальное оперение сохраняется до следующей линьки. В период весенней неполной линьки, как правило, в феврале – марте, сменяется оперение, сохранившееся от гнездового наряда. Основная масса взрослых птиц линяет до осенней миграции – с июля до конца сентября. Первыми у взрослых птиц начинают сменяться кроющие головы и тела. Параллельно выпадают первостепенные маховые, обычно по 2 сразу (1-е и 7-е или 5-е и 10-е), смена их идет в сторону кисти. Такой порядок смены перьев обеспечивает дрофам возможность сохранять хорошие летательные качества в течение круглого года. Из рулевых первыми выпадает центральная пара, что происходит чуть позже смены маховых. При этом самцы начинают линьку несколько раньше самок. Зимняя предбрачная линька взрослых птиц неполная – сменяются кроющие головы, шеи и груди, начинается с конца декабря, но в основном происходит в марте – апреле. Она начинается с задней стороны шеи и «усов» и затем равномерно распространяется на остальные части шеи и груди [389, 486].
Территориальные самцы покидают индивидуальные участки в конце июня, когда самки из их группировок воспитывают птенцов. Сначала они объединяются в самостоятельные небольшие группы, а затем, вероятно, объединяются в большие по размерам стаи с неразмножавшимися в данном полевом сезоне самцами. Таким образом, одиночные самцы в период вождения птенцов на гнездовых территориях не встречаются. Самки с нелетными птенцами держатся в этот период отдельными семейными группами преимущественно на стерне озимых и яровых. В составе каждой из таких стайных структур с интеграционными формами поведения, очевидно, объединяются члены одной территориальной репродуктивной группы [26].
Р
груди
за лопатками (R.)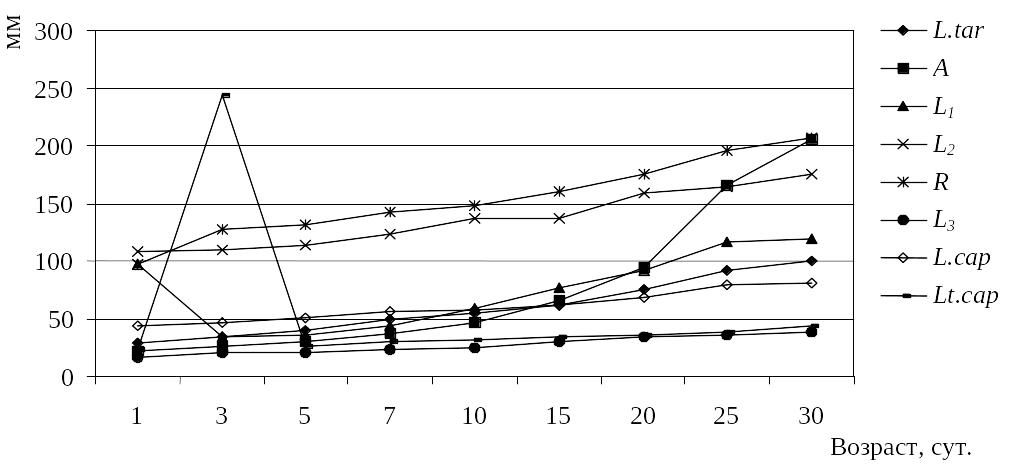 Рис.
22. Ход роста
птенцов дрофы: длины кисти (A),
предплечья (L1),
туловища (L2),
цевки (L.tar),
среднего пальца (L3),
головы (L.cap),
ширины головы (Lt.cap)
и обхвата
Рис.
22. Ход роста
птенцов дрофы: длины кисти (A),
предплечья (L1),
туловища (L2),
цевки (L.tar),
среднего пальца (L3),
головы (L.cap),
ширины головы (Lt.cap)
и обхвата
К
Рис.
23. Соотношение продолжительности
различных стадий репродуктивного
процесса дрофы в Саратовской области
(по оси Y
– число регистраций стадии репродуктивного
процесса):
лупление
птенцов,
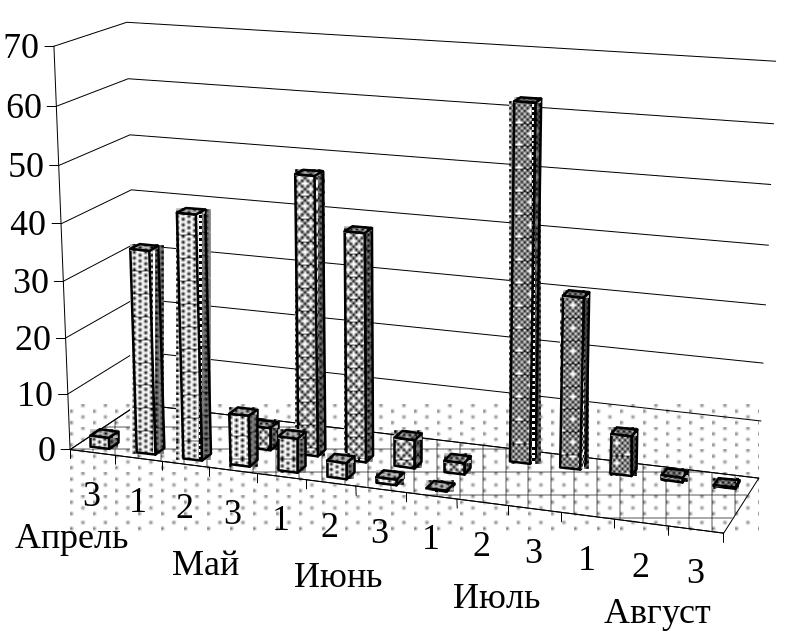
![]() – откладка яиц,
– откладка яиц,
![]() – вы-
– вы-![]() – подъем птенцов на крыло
– подъем птенцов на крыло
В осенний период характерной чертой этологии вида является стремление объединяться в группы. Половозрастная структура изучаемой популяции в это время представлена социальными объединениями нескольких типов: выделены группировки однополых птиц, главным образом самцов в возрасте свыше трех лет и неполовозрелых (как самцов, так и самок) дроф. В категорию одиночных входят птицы обоего пола. Например, в осенний период 1999 г. численность таких агрегаций составляла не более 41 особи, и лишь на отдельных участках наблюдались группировки, состоящие, по-видимому, из неразмножающих птиц (как самцов, так самок) с численностью до 87 особей, ведущих кочевой образ жизни. При появлении всходов озимых культур большая часть дроф концентрируется именно на них. На стерне яровых, ячменя и проса в это время встречаются преимущественно однополые группы самок или самки с выводками. Дрофы активны в течение почти всего светлого времени суток [26].
Известны случаи зимовок дроф в Пугачевском, Питерском и Александровогайском районах [48, 91, 97], Л.А. Лебедева [320] указывала на зимовку 4 особей в 1967 г. в Новоузенском районе. В декабре 1996 г. по 90 особей было зарегистрировано в Энгельсском и Ровенском районах, отдельные птицы отмечались здесь в январе (n = 3) и марте (n = 7) [431]. В январе – феврале 2001 г. две дрофы держались около р. Избалык в сопредельном Павловском районе Ульяновской области. В окрестностях с. Шиковки на поле возле ур. Торновый овраг в тот же период отмечены две пары (устн. сообщ. О.В. Бородина).
Питание. Данные литературы о питании дрофы на севере Н. Поволжья достаточно отрывочны. Известно, например, что в репродуктивный период желудки птиц заполнены, главным образом, зеленой массой, где доминируют листья кермека (Statice Gmelini). Сведения более раннего характера указывают на доминирование в питании дрофы в летний период хлебных побегов и семян [484]. Аналогичные данные приблизительно в этот же период приводит В.Ф. Зуев [485], который сообщает о питании птиц семенами и травой. Более широкий спектр кормов приводится в работе В. Левшина [333], который считает, что «…дрофа, до созревания семян и трав, питается стручками и другой зеленью, а после – исключительно семенами». В осенний рацион вида автор включает молодую рожь, репную траву, молодую репу (Brassica rapa), птенцов жаворонков, куропаток и перепелов, а в зимний – озимь. Конечно же, высказанное мнение о добыче дрофой птенцов птиц характеризуется низкой достоверностью, вместе с тем подобные факты упоминались в литературе и позже.
Кроме того, дрофы поедают большое количество насекомых, среди которых преобладают жесткокрылые, в частности медляки – Blaps [91]. Между тем Р.А. Девишев [330] определяет долю насекомых в пищевом спектре вида в 98%, указывая, что дрофа наиболее часто поедает перелетную саранчу, кузек, свекловичных долгоносиков (Bothynoderes punctiventris). Например, в одном из проанализированных этим автором желудков отмечено 1106 кузек, в другом – 4460 рапсовых листоедов (Entomoscelis adonidis). По данным Б. Лицбарски с соавторами [499], плотность населения членистоногих на участках обитания птенцов дрофы в первые дни их жизни не должна опускаться ниже 9.0 г/м2. При более низких значениях суммарной биомассы беспозвоночных нормальное развитие птенцов нарушается, что приводит к их гибели.
Анализ данных литературы позволяет сделать предварительное заключение о том, что питание дроф в регионе очень разнообразно. При сопоставлении всех имеющихся данных создается впечатление об изменчивости в рационе соотношения животных и растительных компонентов. Безусловно, что в первую очередь это определяется возрастом, полом, временем года, характером гнездовых биотопов и, наконец, индивидуальными особенностями птиц. Следует отметить, что растительная пища постоянно занимает важное место в питании дрофы. Чаще всего это зеленые сочные побеги. Дрофы явно предпочитают их семенам или стеблям, которые, хотя и поедаются, но плохо перевариваются и выделяются с экскрементами в мало изменившемся виде.
Животную пищу дрофы используют в основном в летний период и при повышении ее обилия могут временно полностью переключатся на данные виды кормов. Подобная ситуация возникает, в частности, при массовых вспышках размножения жука кузьки на посевах зерновых, что особенно часто наблюдается в конце июня в некоторых районах саратовского Левобережья. При массовом роении жуков корнегрызов (Risotropys solticialis), в годы массового размножения грызунов – обыкновенной полевки, степной пеструшки, дрофы также в значительной степени переключаются на питание этими видами. На основе полевых наблюдений создается впечатление, что самцы в меньшей степени используют животную пищу. Однако имеющиеся анализы содержимого желудков слишком разнородны, чтобы сделать однозначное заключение о количественном соотношении животной и растительной пищи у половых групп [400].
Наблюдения за взрослыми дрофами в условиях неволи показывают, что при кормлении зернами ячменя, пшеницы, подсолнечника в сухом виде птицы полностью игнорируют данные корма, но охотно поедают распаренные или пророщенные семена. При этом непереваренные зерна значительно чаще отмечаются в помете самок. Многолетние визуальные наблюдения за поведением птиц в природе показывают, что иногда пасущиеся на полях дрофы вырывают подземные части пырея ползучего (Elytrigia repens), луковицы птицемлечников (Ornitoga sp.) и разновидностей лука (Alium sp.).
Наши наблюдения не подтверждают привязанности дроф к водопоям. В условиях саратовского Заволжья, даже на территориях с низким уровнем фактора беспокойства, дрофы не регистрируются вблизи воды. В тех случаях, когда небольшие водоемы сохраняются в центре обширного засеянного поля, где держится несколько десятков дроф, никогда не отмечалось, чтобы птицы подходили к воде. Последнее обстоятельство легко подтверждается на основе анализа следов птиц, которые хорошо выявляются по берегам степных водоемов для других видов (журавлей, чибисов, грачей и даже жаворонков). При осуществлении визуальных наблюдений за дрофами из укрытий отмечено, что птицы могут несколько дней держаться на локальном участке, довольствуясь влагой, содержащейся, очевидно, в сочной растительности, а также росой. Однако в этом случае трудно представить, как обходится без воды самка, насиживающая кладку на пашне, где полностью отсутствует какая-либо растительность. Проводя наблюдения из укрытий, расположенных в 4–5 м от гнезда, можно определенно утверждать, что самка не покидает кладку и не пьет воды. В данном случае можно лишь предположить, что дефицит влаги компенсируется за счет внутренних метаболических процессов, а также жидкости, содержащейся в насекомых, в частности, полевых жужелицах (Carabus arvensis) и других насекомых, которых она добывает в непосредственной близости от гнезда [400].
С целью более детального анализа пищевого спектра дроф нижневолжской популяции обратимся к результатам исследований, проведенных в весенний период 1999 г. в местах скоплений птиц на территории саратовского Заволжья [500]. Изучалось содержимое экскрементов дроф, собранных в непосредственной близости от гнезд, а также содержимое желудков. Всего было обработано 2 желудка дроф, разбившихся о провода ЛЭП (с. Алексашкино Питерского района и с. Ждановка Дергачевского района), и 46 проб экскрементов (с. Светлое Озеро Озинского района – 15, с. Целинный Краснопартизанского района – 9, с. Первомайское Дергачевского района – 22).
При камеральной обработке экскрементов и содержимого желудков установлено, что в весенний период в рационе изучаемого вида доминирует пища растительного происхождения (семена зерновых культур, остатки колосьев, корни и стебли злаков), составляя в выборках от 55% (Дергачевский район) до 90% (Федоровский). В постгнездовой период доля растительности в пище составляет около 64% [362]. Вместе с тем в содержимом желудков весной отмечается большое количество (до 40%) примесей (стекла, фарфора, камней). Животные объекты добывались птицами значительно реже, в питании дрофы были отмечены насекомые из отрядов двукрылых, прямокрылых и жесткокрылых. Последние в пробах экскрементов были представлены жужелицами (золотоямчатой – Carabus clathratus, венгерской – C. pannonicus и хлебной – Zabrus tenebrioides), копром лунным – Copris lunaris, кузькой посевным – Anisoplia segetum, щелкуном черным – Athous niger и усачом подсолнечниковым – Agapanthia dahli. Из прямокрылых птицами добывался только сверчок полевой – Gryllus campestris. Кроме насекомых в экскрементах обнаружены остатки паукообразных [368]. Дрофы, помимо перечисленных объектов питания, потребляли рептилий – прытких ящериц. В серии проб, собранных на территории Краснопартизанского административного района, доля этого вида составила 5% от объема животных остатков [500].
В отношении дрофы в регионе также осуществлено исследование, посвященное анализу питания вида в постгнездовой период. Изучение рациона основано на анализе проб содержимого желудков птиц, а также их экскрементов, собранных с 5 по 25 сентября 2002 г. на убранных полях зерновых культур Краснокутского и Федоровского районов Саратовской области. Для всех проб описывали качественный состав, т.е. наличие тех или иных объектов, прошедших пищеварительный тракт птицы. Затем проводили количественный анализ состава пробы, определяя долю каждого из объектов в единицах объема (% от всех объектов пробы) и его массу. Вегетативные части высших растений, чаще всего в виде листовых пластинок, разделяли на зеленые (вегетирующие) и сухие. Принадлежность остатков животного происхождения определяли по возможности до вида. Всего было обработано 73 пробы экскрементов и содержимое 3 желудков дроф, разбившихся о провода ЛЭП.
Анализ содержимого проб показал, что в пищевом рационе дроф в постгнездовой период пища растительного происхождения (семена зерновых культур, остатки колосьев, стебли растений) явно преобладает над животной (табл. 16). Растительные корма представлены в основном зернами пшеницы, составляя в разных пробах от 22 до 264 экз. (в среднем 114 зерен). Обнаружены также листовые пластинки и плоды сложноцветных (Asteraceae), семена и стебли злаков (Poaceae).
Таблица 16