

Особенности строения половых клеток Строение половых клеток
Строение половых клеток.
Половые клетки, или гаметы, специализированы для полового размножения, поэтому они всегда гаплоидны. В этом заключается их главная особенность. Гаплоидность гамет возникает во время гаметогенеза, т. е. в процессе их образования, и отражает зрелость гамет, их готовность к оплодотворению. Кроме того, сперматозоиды и яйцеклетки отличаются от соматических клеток измененными ядерно-плазменными отношениями. В яйцеклетках по сравнению с соматическими клетками объем цитоплазмы во много раз превышает объем ядер. Яйцеклетки — это обычно самые крупные клетки организма. Примером может служить яйцеклетка птиц. Сперматозоиды, наоборот, имеют очень мелкие размеры. В них почти нет цитоплазмы. В половых клетках по сравнению с соматическими резко снижен обмен веществ. Процессы ассимиляции и диссимиляции в них минимальны.
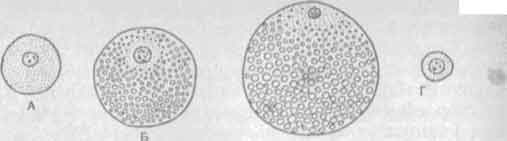
Рис. 30. Строение яйцеклеток различных типов.
Половые клетки характеризуются особым строением, отражающим их функциональное назначение. Гак, яйцеклетки не способны к самостоятельному перемещению. Большие размеры яйцеклеток объясняются накоплением в них питательных веществ. Такие питательные вещества, как жир, гликоген и особые белки — фосфопротенны, используются на начальных стадиях развития зародыша. Фосфопротеины составляют основную часть запаса. Они получили название желтка. Количество желтка и его распределение в яйцеклетках различных видов организмов неодинаковы. На примере хордовых животных четко прослеживаются исторические видоизменения размеров яйцеклеток. Чем сложнее устроены организмы и чем длительнее у них эмбриональный период, тем больше желтка накапливается в яйцеклетках. Исключение составляют яйцеклетки плацентарных млекопитающих, которые практически не содержат запаса вещества в связи с переходом к внутриутробному развитию зародышей. Это как бы вторичное уменьшение количества желтка в яйцеклетках.
Яйцеклетки животных классифицируют по содержанию в них желтка и по тому, как этот желток распределен (рис. 30). Изолецитальные яйцеклетки ланцетника имеют небольшие размеры, желтка в них мало, распределен он равномерно. Ядро находится почти в центре. Подобные яйцеклетки имеются и у беспозвоночных животных, таких как черви, моллюски, иглокожие. Умеренно телолецитальные яйцеклетки земноводных и осетровых рыб содержат значительно больше желтка, размеры их достигают 1,5—2 мм. Основная часть желтка сконцентрирована на одном из полюсов, называемом вегетативным. На противоположном— анимальном — полюсе желтка мало. Здесь находится ядро яйцеклетки. Таким образом, для телолецитальной яйцеклетки характерно неравномерное распределение желтка. Резко телолецитальные яйцеклетки пресмыкающихся, птиц, некоторых рыб и головоногих моллюсков содержат очень много желтка. Размеры их достигают 10—15 мм и больше.
Распределение желтка еще более неравномерное. На анимальном полюсе находится зародышевый диск с ядром и активной цитоплазмой, а вся остальная часть клетки занята желтком. Алецитальными называются яйцеклетки, в которых очень мало желтка, размеры их малы. Яйцеклетка женщины имеет диаметр 100—300 мкм. Количество и распределение желтка в яйцеклетке определяют характер дробления, гаструляции и вообще тип эмбрионального развития.
Сперматозоиды — очень мелкие и подвижные клетки. Например, величина сперматозоидов человека — 50—70 мкм, крокодила — 20 мкм. Главная функция сперматозоида — внести при оплодотворении свой гаплоидный набор хромосом в яйцеклетку. Строение сперматозоида соответствует его функциям. Чаще всего он имеет головку и жгутик (хвост), соединенные промежуточным отделом — шейкой (рис. 31). В головке размещаются ядро и особая структура — акросома. Акросома содержит ферменты, обеспечивающие проход ядра сперматозоида в яйцеклетку при оплодотворении. В промежуточном отделе лежат центриоли клеточного центра и митохондрии. Из центриоли отходит жгутик, обеспечивающий подвижность. Встречаются и безжгутиковые сперматозоиды.

Рис. 31. Строение жгутикового сперматозоида.
д _ головка; Б — промежуточный отдел; В — жгутик; I — акросома; 2 — ядро; 3 — центриоли; 4 — метохондрии жгутика; 5 — осевая нить.
36
Образование половых клеток. Мейоз. Фазы мейоза Мейоз (от греч. meiosis — уменьшение) или редукционное деление клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать сгаметогенезом — образованием специализированных половых клеток, или гамет, из недифференцированных стволовых.
С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса.
В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма.
Этот же механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет. Определенные ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации).
Содержание [убрать]
|