

Классификация
Вторичные посредники классифицируют по растворимости в воде и размеру молекулы
Гидрофобные молекулы — нерастворимые в воде молекулы, например, диацилглицерол, инозитолтрифосфат, фосфатидилинозитолы — связаны с мембраной и могут диссоциировать в околомембранное пространство, где регулируют активность белков, связанных с мембраной;
Гидрофильные молекулы — водорастворимые молекулы и ионы, например, цАМФ, цГМФ, Ca2+ — находятся в цитозоле;
Газы — оксид азота (NO), оксид углерода (CO) — могут переходить из цитозоля в межклеточную среду через клеточные мембраны.
2. Нейробиологическая концепция «командного нейрона».
Процедуру классификации на нейронном уровне можно представить на основе введения в модель следующего слоя обработки информации — командных нейронов. Командный нейрон — это формальный нейрон со многими входами и многими выходами, выполняющий функцию генерации внешней реакции. Выходами командного нейрона являются возбуждения, ветвящиеся по аксонным терминалям и образующие синаптические контакты на мотонейронах. Единичное возбуждение командного нейрона, проходя через разные по эффективности синаптические контакты с мотонейронами, образует вектор возбуждения, вовлекающий в реакцию целый ансамбль мотонейронов, которые, в свою очередь, будучи связанными с мышечными единицами, формируют двигательный аккорд — целостный поведенческий акт или его отдельный фрагмент. В частном случае отдельный командный нейрон может быть ответственным за генерацию речевой реакции. Процедура классификации на уровне командных нейронов реализуется при участии нейронов-детекторов. Потенциально множество селективных нейронов-детекторов подключены параллельно ко всем командным нейронам. Связи детекторов с командными нейронами пластичны, изменяясь в процессе обучения. Процесс обучения происходит в результате сочетания внешнего сигнала с разрядом данного командного нейрона. Те синаптические связи детекторов с командным нейроном, за активацией которых следует разряд данного командного нейрона, усиливаются. Когда связи между некоторыми детекторами с данными командными нейронами возрастают в такой степени, что возбуждение командного нейрона достигает порога генерации спайков, сигнал вызовет разряд командного нейрона и связанную с ним поведенческую реакцию. Те синаптические контакты между детекторами и командным нейроном, которые подвергаются активации при действии сигнала, но при этом их активация не сопровождается последующим разрядом командного нейрона, ослабевают. Таким образом, на данном командном нейроне формируется матрица высокоэффективных и низкоэффективных синапсов, через которые к нему поступают сигналы от нейронов-детекторов. Внешние сигналы, воздействуя на те детекторы, которые оканчиваются на командном нейроне высокоэффективными синапсами, будут селективно вызывать реакцию данного командного нейрона и ту форму поведения, которая этим командным нейроном реализуется по механизму условного рефлекса. Подкрепляющие воздействия, всегда приводящие к разряду командного нейрона, поступают к нему по непластичным возбуждающим синапсам, вызывая разряд командного нейрона и связанное с ним поведение по механизму безусловного рефлекса. Командные нейроны принимают участие в генерации речевых реакций. Пусть имеется некоторый набор сигналов, каждый из которых вызывает максимум возбуждения на разных нейронах-детекторах. В процессе обучения каждый сигнал через нейроны-детекторы селективно связывается с определенным командным нейроном речевой реакции. В результате внешний сигнал вызывает специфически связанную с ним в процессе обучения реакцию, реализуя таким образом простейшую форму классификации сигналов.
Билет16
1.Регуляция полового и родительского поведения.
У млекопитающих гипофиз секретирует гонадотропные гормоны, которые оказывают регулирующее влияние на различные физиологические процессы, имеющие отношение к размножению. Наибольший эффект гонадотропные гормоны оказывают на функционирование половых желез. Гормоны мужских и женских половых желез (андрогены и эстрогены), действуя на ЦНС, активируют половое поведение. В организме взрослой женщины наблюдаются периодические изменения различных функций, связанные с повторяющимися процессами роста и созревания фолликулов, овуляции и образования желтых тел. Созревание фолликулов сопровождается высоким уровнем секреции половых гормонов. В этот период яйцеклетка освобождается из фолликула (овуляция) и попадает в половые пути.. Важную роль здесь играет действие эстрогена на гипоталамус. Удаление у крысы яичников вызывает снижение и прекращение половой активности. Введение эстрогена в гипоталамус овариэктомированных (удалены яичники) самок сопровождается всем комплексом проявлений эструса. Наибольшее значение для организации полового поведения самок, по-видимому, имеет гипоталамус. Его разрушение приводит к потере половой активности. При локальных повреждениях в различных областях гипоталамуса можно заблокировать либо поведение, характерное для эструса, либо секрецию гонадотропного гормона. В эмбриональном или раннем постнатальном периоде развития (в зависимости от вида животного) происходит половая дифференцировка головного мозга по отношению к регуляции секреции гонадотропных гормонов гипофиза, реактивности к половым гормонам, поведению, росту и др. Половые различия мозга определяются разными структурами и прежде всего медиальным гипоталамусом. Важным фактором указанной дифференцировки является действие на мозг в «критический период» развития организма половых гормонов. До этого мозг независимо от генетического пола эмбриона или новорожденного имеет женский тип. Под влиянием андрогена, вырабатываемого семенниками, мозг маскулинизируется, т. е. приобретает мужской тип. Большая часть вегетативных и двигательных реакций, составляющих половой акт, интегрируется на спинальном уровне в поясничных и крестцовых сегментах. После перерезки спинного мозга путем стимуляции половых органов можно вызвать эрекцию и даже эякуляцию. Гипоталамус участвует в регуляции половой активности у самца. Миндалина также включена в систему контроля полового поведения. Возможно, в норме миндалина оказывает тормозное влияние на половое поведение. Значение коры больших полушарий в организации полового поведения в ходе эволюции млекопитающих возрастает.
2.Роль ионных насосов в формировании ПП.
Потенциа́л поко́я (ПП) - мембранный потенциал возбудимой клетки в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 мВ[1]. У нейронов и нервных волокон обычно составляет -70 мВ. Измеряется изнутри клетки.
В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация внутри и вне клетки постепенно должна выравниваться. Однако, несмотря на постоянную диффузию ионов (утечка ионов), ПП клеток остается на одном уровне. Следовательно, кроме собственно ионных механизмов формирования ПП, связанных с различной проницаемостью клеточной мембраны и диффузией ионов, имеется активный механизм поддержания градиентов концентрации различных ионов внутри и вне клетки. Таким механизмом являются ионные насосы, в частности Na/К-насос (помпа).Ионный насос - это транспортная система, обеспечивающая перенос иона с непосредственной затратой энергии вопреки концентрационному и электрическому градиента. Если ионов К+ на наружной стороне мембраны нет, то работа насоса блокируется, перенос ионов Nа+ из клетки в этом случае падает, составляя примерно 30% от нормального уровня. Накопление Na+ в клетке стимулирует работу Nа/К-насоса, уменьшение Na+ в клетке снижает его активность, поскольку снижается вероятность контакта ионов с соответствующим переносчиком. В результате сопряженного транспорта ионов Na+ и К+ поддерживается постоянная разность концентраций этих ионов внутри и вне клетки. Одна молекула АТФ обеспечивает один цикл работы Ка/К- насоса - перенос 3 ионов Nа+ за пределы клетки и 2 ионов К+ внутрь клетки. Асимметричный перенос ионов Na/K-насосом поддерживает избыток положительно заряженных частиц на наружной поверхности клеточной мембраны и отрицательных зарядов внутри клетки, что позволяет считать Na/K-насос структурой электрогенной, дополнительно увеличивающей ПП примерно на 5-10 мВ. Данный факт свидетельствует о том, что решающим фактором в формировании ПП является селективная проницаемость клеточной мембраны для разных ионов. мембрана малопроницаема для Nа+,поэтому диффузия в обратном направлении происходит очень медленно. Для К+ мембрана клеток в покое более проницаема, соответственно К+ диффундирует наружу гораздо быстрее. Ионный насос поддерживает разность концентраций ионов Nа+ и К+ вне и внутри клетки. Нормальная величина ПП является необходимым условием возникновения процесса возбуждения клетки, т.е. возникновения и распространения ПД, инициирующего специфическую деятельность клетки.
Билет17
1.Рецепторы. Рецепторный и генераторный потенциал.
Рецепторы представляют собой специализированные образования, воспринимающие определенные виды раздражений.
Рецепторы обладают наибольшей чувствительностью к адекватным для них раздражениям. Рецепторы делят на четыре группы: механо-, термо-, хемо- и фоторецепторы. Каждую группу подразделяют на более узкие диапазоны рецепции. Например, зрительные рецепторы делятся на воспринимающие освещенность, цвет, слуховые — определенный тон, вкусовые — определенные вкусовые раздражения (соленое, сладкое, горькое) и т. д.
Рецепторный потенциал возникает при раздражении рецептора как результат деполяризации и повышения проводимости участка его мембраны, который называется рецептивным. Рецептивный участок мембраны имеет специфические свойства, в том числе биохимические, отличающие его от мембраны тела и аксона.
Возникший в рецептивных участках мембраны рецепторный потенциал электротонически распространяется на аксонный холмик рецепторного нейрона, где возникает генераторный потенциал. Возникновение генераторного потенциала в области аксонного холмика объясняется тем, что этот участок нейрона имеет более низкие пороги возбуждения и потенциал действия в нем развивается раньше, чем в других частях мембраны нейрона. Чем выше генераторный потенциал, тем интенсивнее частота разрядов распространяющегося потенциала действия от аксона к другим отделам нервной системы. Следовательно, частота разрядов рецепторного нейрона зависит от амплитуды генераторного потенциала.
Рецепторные нейроны различаются по скорости уменьшения их реакции (адаптации) на длящуюся стимуляцию. Рецепторные нейроны, медленно адаптирующиеся к раздражению, т. е. длительное время генерирующие потенциалы действия, называются тоническими. Рецепторы, быстро и коротко реагирующие на стимуляцию группой импульсов, называются физическими.
Таким образом, реакция рецепторного нейрона, предназначенного для передачи информации из области восприятия, имеет 5 стадий: 1) преобразование сигнала внешнего раздражения; 2) генерация рецепторного потенциала; 3) распространение рецепторного потенциала по нейрону; 4) возникновение генераторного потенциала; 5) генерация нервного импульса.
2. Нервный центр. Свойства нервных центров.
Нервным центром (НЦ) называется совокупность нейронов в различных отделах ЦНС, обеспечивающих регуляцию какой-либо функции организма. Например, бульбарный дыхательный центр.
Для проведения возбуждения через нервные центры характерны следующие особенности:
1. Одностороннее проведение. Оно идет от афферентного, через вставочный, к эфферентному нейрону. Это обусловлено наличием межнейронных синапсов.
2. Центральная задержка проведение возбуждения. Т.е. по НЦ возбуждение идет значительно медленнее, чем по нервному волокну. Это объясняется синаптической задержкой. Так как больше всего синапсов в центральном звене рефлекторной дуги, там скорость проведения наименьшая. Исходя из этого, время рефлекса, это время от начала воздействия раздражителя до появления ответной реакции. Чем длительнее центральная задержка, тем больше время рефлекса. Вместе с тем оно зависит от силы раздражителя. Чем она больше, тем время рефлекса короче и наоборот. Это объясняется явлением суммации возбуждений в синапсах. Кроме того, оно определяется и функциональным состоянием ЦНС. Например, при утомлении НЦ длительность рефлекторной реакции увеличивается.
3. Пространственная и временная суммация. Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда ВПСП. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторов нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются и в мембране нейрона генерируется распространяющийся ПД.
4. Трансформация ритма возбуждения – изменение частоты нервных импульсов при прохождении через нервный центр. Частота может понижаться или повышаться. Например, повышающая трансформация (увеличение частоты) обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне (рис). Второе, генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких ВПСП и возникновением одного ПД в нейроне.
5. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы,. выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
6. Последействие, это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
7. Тонус нервных центров – состояние постоянной повышенной активности. Он обусловлен постоянным поступлением к НЦ нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например проявлением тонуса соответствующих центров является тонус определенной группы мышц.
8. Автоматия или спонтанная активность нервных центров. Периодическая или постоянная генерация нейронами нервных импульсов, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов.
9. Пластичность нервных центров. Это их способность изменять функциональные свойства. При этом центр приобретает возможность выполнять новые функции или восстанавливать старые после повреждения. В основе пластичности Н.Ц. лежит пластичность синапсов и мембран нейронов, которые могут изменять свою молекулярную структуру.
10. Низкая физиологическая лабильность и быстрая утомляемость. Н.Ц. могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов.
Билет 18
1.Ноцицептивная система.
Ноцицепция (синонимы: ноциперцепция, физиологическая боль) — это активность в афферентных (приносящих) нервных волокнах периферийной и центральной нервной системы, возбуждаемая разнообразными стимулами, обладающими «повреждающей» интенсивностью. Данная активность генерируется ноцицепторами, или по-другому рецепторами боли, которые могут отслеживать механические, тепловые или химические воздействия, превышающие генетически установленный порог обычного восприятия. Получив повреждающий стимул, ноцицептор передаёт сигнал через спинной мозг далее в головной мозг. Ноцицепция сопровождается также самыми разнообразными проявлениями и может служить для возникновения опыта боли у живых существ. Передача болевых сигналов в центральной нервной системеСуществуют два направления, по которым происходит передача сигналов в головной мозг. Это неоспинобугорный путь (для быстрой боли) и палеоспинобугорный путь (для медленной боли). Неоспиннобугорный путь. Быстрая боль направляется через Aδ-волокна, которые заканчиваются в сегменте I заднего рога спинного мозга. Здесь вторые по порядку в этом пути, передаточные нейроны вступают в дальнейший контакт и поднимают сигнал через позвоночные столбы. Эти волокна затем пересылают сигнал в вентролатеральные ядра таламуса. Оттуда третьи нейроны связываются с соматосенсорными полями коры головного мозга. Быстрая боль легко локализуется, если Aδ-волокна стимулируются совместно с тактильными рецепторами. Палеоспиннобугорный путь. Медленная боль передаётся через медленный тип С-волокон к сегментам II и III заднего рога спинного мозга, также именуемым студенистым веществом. Здесь вторые по порядку в этом пути нейроны контактируют и пересылают сигнал в сегмент V заднего рога спинного мозга. Затем третьи нейроны присоединяются к волокнам быстрого пути, и направляются через переднелатеральный путь. Эти нейроны широко распространены в стволе мозга, одна десятая часть которых завершается в таламусе, а остальные — в продолговатом мозге, мосту и среднем мозгу. Медленную боль сложно локализовать.
2.Серотонин: периферические и центральные эффекты. 5-НТ-рецепторы, их разнообразие и функции.
Серотонин, 5-окситриптамин, биологически активное вещество, содержащееся в крови и тканях животных и человека; является медиатором нервной системы как на периферии, так и в нервных центрах (главным образом в гипоталамусе). Впервые выделен
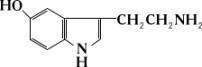
и получен в кристаллическом виде из сыворотки быка (1947). В организме С. синтезируется из аминокислоты триптофана, у человека и большинства позвоночных — главным образом в клетках слизистой оболочки тонкого кишечника, а также в поджелудочной железе и центральной нервной системе. В тканях С. превращается в 5-оксииндолилуксусную кислоту, которая выводится из организма почками. В процессах обмена С. большую роль играет фермент моноаминоксидаза.
Рецепторы серотонина представлены как метаботропными, так и ионотропными. Всего насчитывается семь типов таких рецепторов, 5-HT 1-7, причем 5-НТ 3 — ионотропные, остальные — метаботропные, семидоменные, связанные с G-белками. Установлено сходство метаботропных 5-HT рецепторов с рецепторами норадреналина.
5-HT 1 тип, насчитывающий несколько подтипов: 1А-E, которые могут быть как пре- так и постсинаптическими, подавляет аденилатциклазу; 5-НТ 4 и 7 — стимулируют; 5-HT 2, насчитывающий несколько подтипов: 2А-C, которые могут быть только постсинаптическими, активирует инозитолтрифосфат. 5-HT 5A подтип также подавляет аденилатциклазу.[1]
Для некоторых типов рецепторов обнаружены эндогенные лиганды, помимо серотонина. Это, например, 5HT-модулин (Leu-Ser-Ala-Leu), лиганд 1B и 1D пресинаптических рецепторов, индуктор тревожности и стресса.
Структура серотонина имеет сходство со структурой психоактивного вещества ЛСД. ЛСД действует как агонист некоторых 5-HT рецепторов и ингибирует обратный захват серотонина, увеличивая его содержание.