

1.5.Холинэргическая проводимость
Медиатором холинэргической проводимости служит ацетилхолин (АХ). Он синтезируется в концевых пластинах нерва под действием холин-ацетилтрансферазы, которая транспортируется в концевые пластинки эндоплазматического ретикулума:
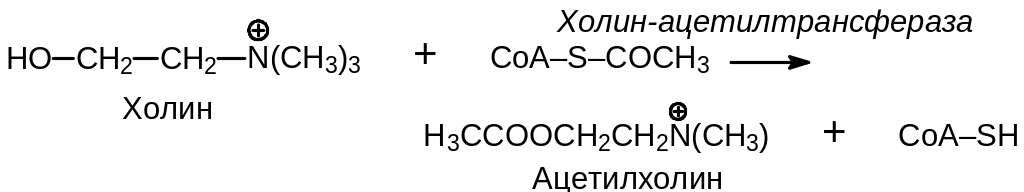
Синтезированный ацетилхолин хранится в синаптических пузырьках. Пузырьки имеют диаметр около 50 нм и содержат примерно 4·104 молекул ацетилхолина (концентрация 0,5 М). Высвобождение ацетилхолина из пузырьков происходит порциями в ответ на возбуждение, передаваемое от аксона. Один импульс вызывает выделение приблизительно 100–200 порций медиатора. Запаса ацетилхолина хватает для 2500-5000 импульсов. После исчерпания его запаса мембраны пузырьков повторно заполняются вновь синтезированными молекулами АХ. Холин, получающийся при расщеплении ацетилхолина, используется для его повторного синтеза.
После освобождения в синаптическую щель, ацетилхолин пересекает ее посредством диффузии. Рецепторные белки (R) находятся в противоположной постсинаптической мембране. Белки-рецепторы ацетилхолина бывают двух типов: мускаринового (медленного) и никотинового (быстрого). Как показывают их названия, эти белки активируются соответственно мускарином и никотином. Их роль заключается в передаче импульса постсинаптической мембране. Этот процесс оказывается возможным благодаря увеличению проницаемости мембраны для Na+ и К+. В случае рецепторов; мускаринового типа в процессе участвует cGMP, который синтезируется из GTP под действием гуанилатциклазы.
Наряду с активаторами рецепторных белков известны и их ингибиторы: для рецепторов мускаринового типа это атропин, а для рецепторов никотинового типа — сукцинилхолин, D-тубокурарин (яд для наконечников стрел) и -бунгаротоксин (змеиный яд). Число рецепторов в нервно-мышечном синапсе было определено с помощью змеиного токсина и оказалось равным 13000 на квадратный микрон. Избыток ацетилхолина в районе синаптической щели вызывал бы постоянную передачу импульсов, но так как это нежелательно, его избыток разлагается под действием специфической гидролазы, ацетилхолинэстеразы. Гидролаза расщепляет ацетилхолин до ацетата и холина, которые затем транспортируются обратно в нервные окончания и используются для ресинтеза.
Карбаматы и метансульфонаты обладают ингибирующим действием на ацетилхолинэстеразу. Высокая токсичность органических производных фосфора (например, диизопропилфторфосфата) объясняется образованием прочной связи с ацетилхолинэстеразой и блокированием ферментативного гидролиза избытка ацетилхолина.
Одно из наиболее ядовитых соединений, ботулотоксин, блокирует выход ацетилхолина из пузырьков.
1.6.Адренэргическая передача
Медиатором адренэргической передачи является норадреналин. Он синтезируется в нервных концевых пластинках в результате многостадийного процесса (Рис. 10 .5).
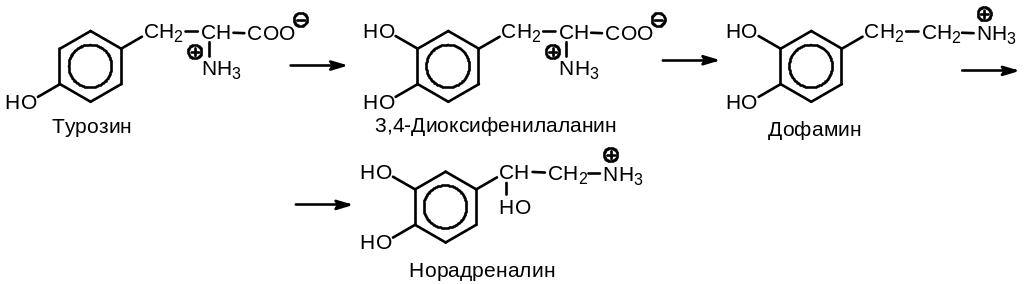
Рис. 10.5. Синтез норадреналина
Гидроксилирование L-тирозина с образованием 3,4-диоксифенилаланина (ДОФА) катализируется тирозин-3-гидроксилазой в присутствии тетрагидроптеридина и NADPH. Донором ОН-группы служит кислород. ДОФА декарбоксилируется ДОФА-декарбоксилазой в присутствии пиридоксальфосфата. Норадреналин образуется из получающегося промежуточного продукта в результате еще одной реакции декарбоксилирования, катализируемой диоксифенилэтиламин--гидроксилазой в присутствии аскорбиновой кислоты и Са2+. Синтезированные молекулы норадреналина хранятся в синаптических пузырьках диаметром 200 нм.
Прохождение импульса вдоль аксона начинается с освобождения медиатора. Последний выходит в синаптическую щель, диффундирует через нее и присоединяется к рецептору в постсинаптической мембране. Существуют два типа рецепторов — и . Передача импульсов -адренэргическим рецептором связана с cGMP. Его агонистом является фенилэфрин, а антагонистами — дибенамин и фентоламин (регитин). Передача импульсов -адрен-эргическим рецептором связана с сАМР. Его агонистами являются адреналин и изопротеренол, а антагонистом — пропранолол. Небольшое количество норадреналина диффундирует из пузырьков без дополнительных воздействий. Эта нежелательная утечка устраняется под действием моноаминоксидазы, фермента, требующего NAD или FAD, которая превращает норадреналин в 3,4-диокси-фенилуксусную кислоту. Избыток норадреналина в синаптической щели инактивируется катехол-О-метилтрансферазой, требующей для своей работы S-аденозилметионин. Таким образом, норадреналин превращается в 3-О-метилнорадреналин (норметанэфрин), который далее окисляется в печени под действием моноаминоксидазы до З-метокси-4-оксиминдальной (ванилилминдальной) кислоты. Эта кислота выводится с мочой (Рис. 10 .6).
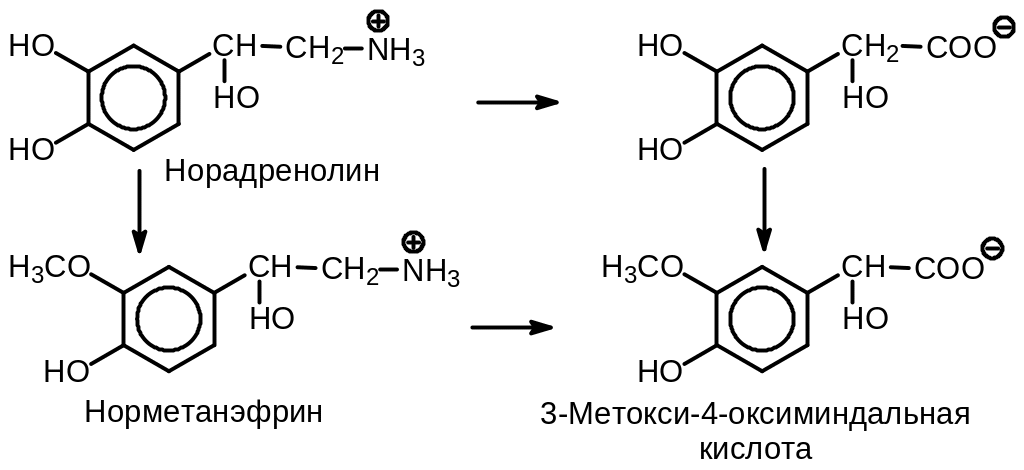
Рис. 10.6. Превращения норадреналина
Рис.
10.7.
Строение
адреналина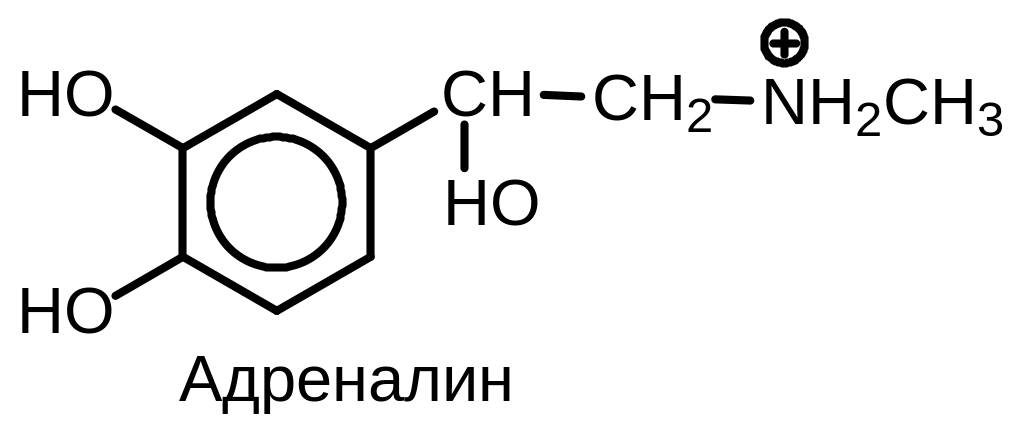
Под действием катехол-О-метилтрансферазы адреналин превращается в 3-О-метиладреналин (метанэфрин), а затем под действием моноаминоксидазы в ванилилминдальную кислоту.