1.1.Репликация днк
Основной функцией матричного синтеза ДНК, катализируемого ДНК-зависимыми ДНК-полимеразами, является удвоение ДНК при делении клетки. Химическая реакция заключается в переносе дезоксирибонуклеотида от дезоксирибонуклеозид-5-трифосфата на 3-гидроксил растущей цепи ДНК (Рис. 6 .1).
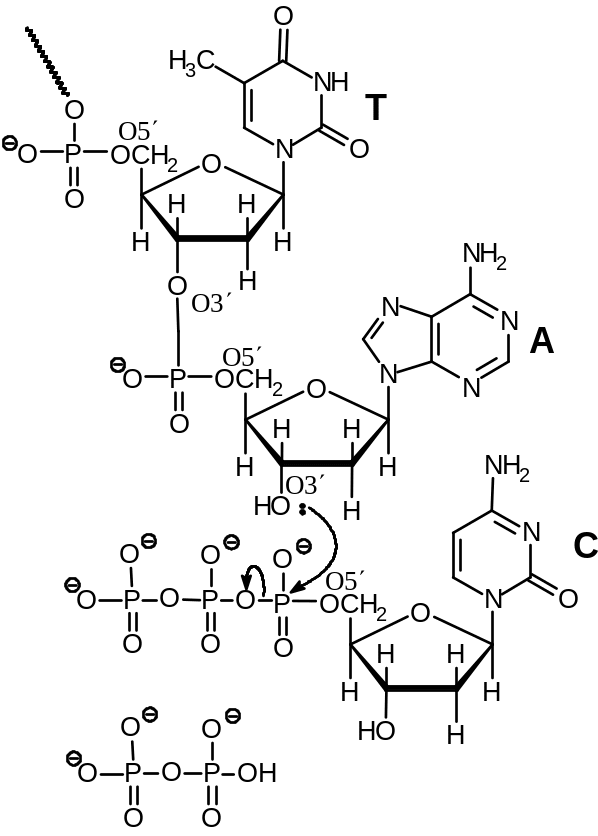
Рис. 6.1. Химическая реакция, катализируемая ДНК-полимеразами.
Использование в качестве субстратов дезоксирибонуклеозид-5-трифосфатов делает синтез полинуклеотида термодинамически выгодным, поскольку он сопрягается с разрывом богатой энергией ангидридной пирофосфатной связи. Необходимая энергия запасается в предыдущих нематричных стадиях синтеза дезоксирибонуклеозид-5-трифосфатов (Рис. 6 .2). Видно, что при этом затрачивается энергия двух молекул АТР.
В клетках как прокариот, так и эукариот имеется несколько различных ДНК-полимераз. В частности для E. coli известно три ДНК-полимеразы: I, II и III. Функции ДНК-полимеразы II мало изучены и она не является жизненно необходимым ферментом. ДНК-полимераза I участвует в процессах репарации. ДНК-полимераза III состоит из семи субъединиц (суммарная молекулярная масса около 380000). Именно она является ферментом репарации.
У эукариот достоверно установлено наличие трех ДНК-полимераз. В репликации ДНК хромосом участвует ДНК-полимераза . ДНК-полимераза катализирует процессы репарации, а ДНК-полимераза — процессы репликации ДНК митохондрий. Важно отметить, что для начала синтеза ДНК-полимеразой необходимо наличие связанного с ДНК короткого олигонуклеотида, называемого затравкой или, чаще, праймером.
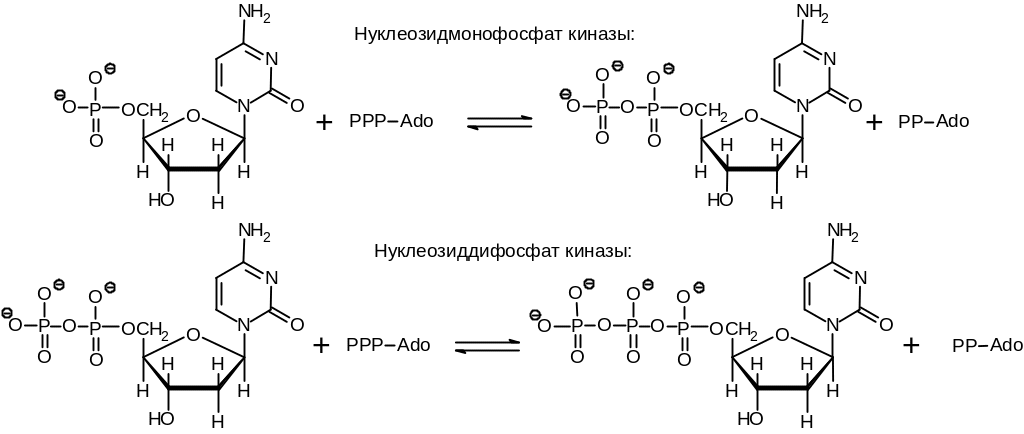
Рис. 6.2. Синтез дезоксирибонуклеозид-5-трифосфатов
Репликация двуцепочечной ДНК в клетке является сложным, далеко не во всех деталях исследованным процессом. Она начинается, как правило, не с концов ДНК, а в строго определенных участках ori (сокращение от термина origin of replication) и распространяется в обе стороны от этого участка. Затем, когда репликация одной их нитей достигает конца молекулы, структура принимает Y-образную форму (Рис. 6 .3). Участок, примыкающий к точке разветвления, в котором и происходят основные события репликации, называют репликативной вилкой (Рис. 6 .3).
Молекула ДНК представляет собой плотно скрученную двойную спираль, причем кодирующие основания находятся внутри этой спирали. Таким образом, для считывания информации цепи должны быть разведены хотя бы на коротком участке. Кроме того, при движении репликативной вилки (Рис. 6 .3) нереплицированная часть ДНК должна раскручиваться. Очевидно, что вращение ДНК как единого целого физически невозможно (отметим, что для обеспечения наблюдаемой скорости репликации ДНК должна вращаться со скоростью 4500 оборотов в мин). Как же решается эта проблема в клетке?
Раскручивание двойной спирали ДНК и удержание двух цепей на некотором расстоянии друг от друга осуществляется с помощью нескольких специальных белков. Ферменты, называемые геликазами, расплетают короткие участки ДНК непосредственно перед репликационной вилкой. По каждой из цепей движется своя геликаза. Та, которая движется в направлении от 3- к 5-концу, называется геликазой или Rep-белком, а та, что движется от 5- к 3-концу, — геликазой II. Направленное перемещение требует затрат энергии и каждый акт перемещения сопровождается гидролизом молекулы АТР. Таким образом, на расплетение одной пары оснований затрачивается энергия двух молекул АТР. Как только небольшой участок ДНК оказывается расплетенным, а каждой из цепей присоединяется несколько молекул ДНК-связывающего белка (ssb-белок от английского термина Single Strand Binding) , которые препятствуют образованию комплементарных пар и обратному воссоединению цепей.
Рис.
6.3. Схема
основных процессов, протекающих в
репликативной вилке.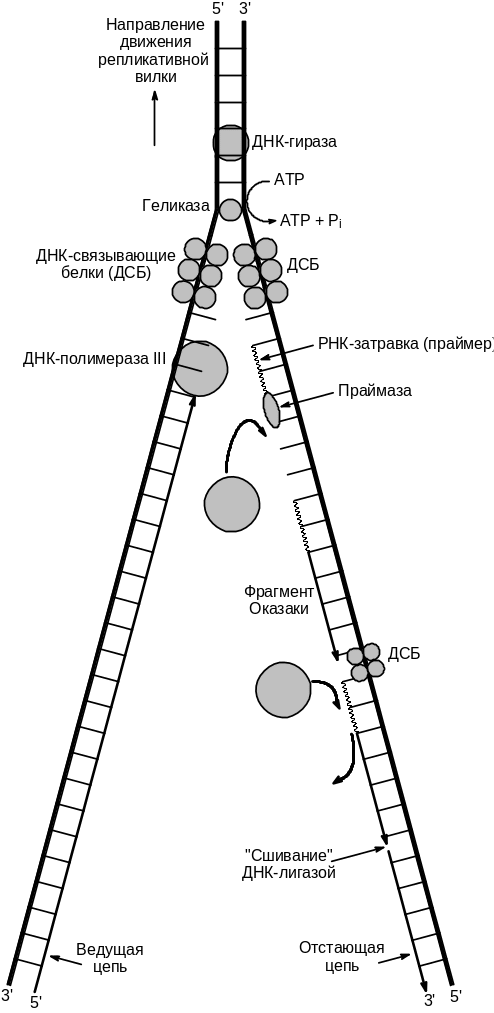
Удаление праймера происходит по двум механизмам. В некоторых случаях это делает сама ДНК-полимераза, которая кроме 35‑синтетазной активности обладает так же 53-эндонуклеазной активностью. В других случаях рибонуклеотидный праймер удаляется специальным ферментом — РНКазой Н. И в том и в другом случае образовавшаяся после удаления праймера брешь застраивается ДНК-полимеразой. И, наконец, вновь синтезированный фрагмент, состоящий уже дезоксирибонуклеотидов, объединяется с цепью ДНК специальным ферментом — ДНК-лигазой.
Развертывание участка двуцепочечной спирали ДНК-гиразами не создавало бы никаких затруднений, если бы концы ДНК были свободными. Однако ДНК в хромосоме прочно связана с белками (гистонами) и плотно уложена и не может вращаться как единое целое (строение этого комплекса мы кратко рассмотрим чуть ниже). Поэтому расплетание на некотором участке приводит к большей закрученности нерасплетенной спирали, т.е. к появлению суперспирализации. Поскольку супервитки совпадают с направлением закручивания исходной спирали, их называют положительными. Суперспирализация порождает напряжения в спирали и в конце концов может воспрепятствовать репликации. Что бы этого не случились, необходимо либо постоянно «сбрасывать» положительные супервитки, либо заранее, до действия гиразы вводить в ДНК отрицательные супервитки. Эту функцию выполняет фермент называемый ДНК-топоизомеразой. Механизм введения отрицательных супервитков заключается в введении разрыва в одну из цепей, поворота одного из концов вокруг целой цепи и восстановления разорванной связи. Процесс сопровождается гидролизом АТР.
Рис.
6.4. Электронная
микрофотография хромосомы 12 человека.
В волокнах хроматина ДНК очень прочно связана с белками, называемыми гистонами. Гистоны содержат много положительно заряженных аминокислотных остатков (лизинов и аргининов) и именно электростатическое притяжение обеспечивает прочное связывание этих белков с ДНК.
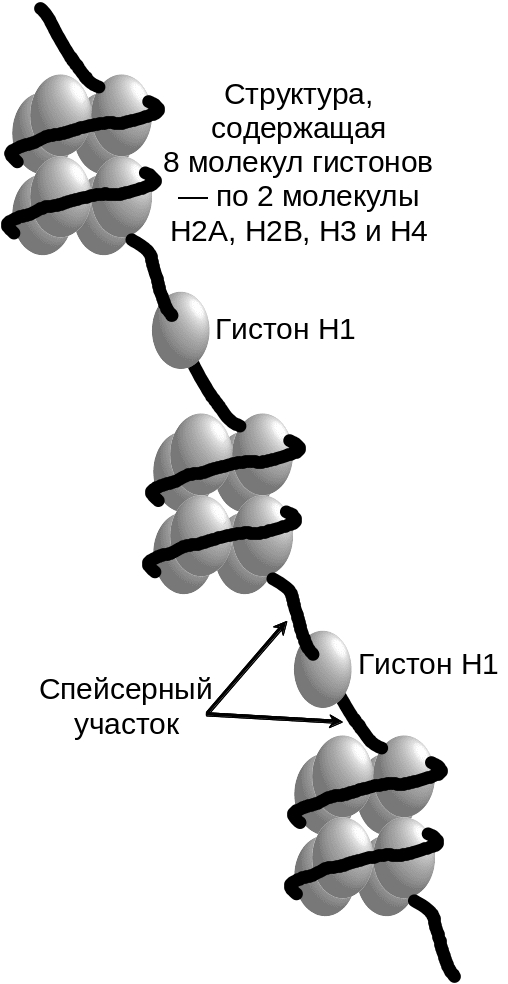
Хроматиновые волокна напоминают по внешнему виду нитку бус (Рис. 6 .5). Повторяющиеся, похожие на бусинки структуры носят название нуклеосом.
Рис.
6.5. Строение
волокон хроматина. Комплекс, состоящий
из 8 молекул гистонов, обвитый двумя
витками ДНК называется нуклеосомой.