

Окисление липидов в тканевом липолизе
Продукты гидролиза липидов — жирные кислоты и глицерин — способны окисляться в тканях до СО2 и Н2О. Освобождающаяся при этом химическая энергия частично аккумулируется в богатых энергией фосфатных связях АТФ, частично рассеивается в виде тепла. Так как химическая природа жирных кислот и глицерина различна, различны и пути их окисления.
Превращение глицерина
Окисление глицерина начинается с его фосфорилирования; при взаимодействии глицерина с АТФ образуется фосфоглицерин.
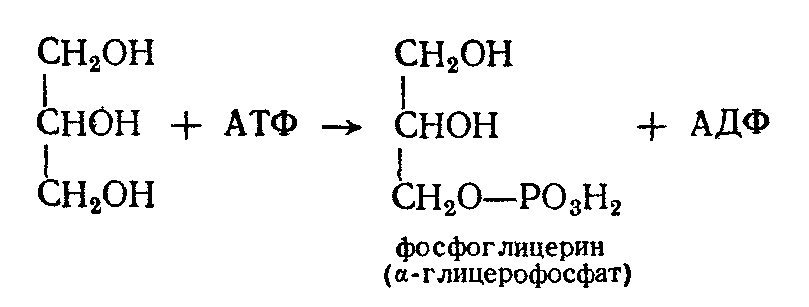
Затем он дегидрогенируется, образуя фосфоглицериновый альдегид:
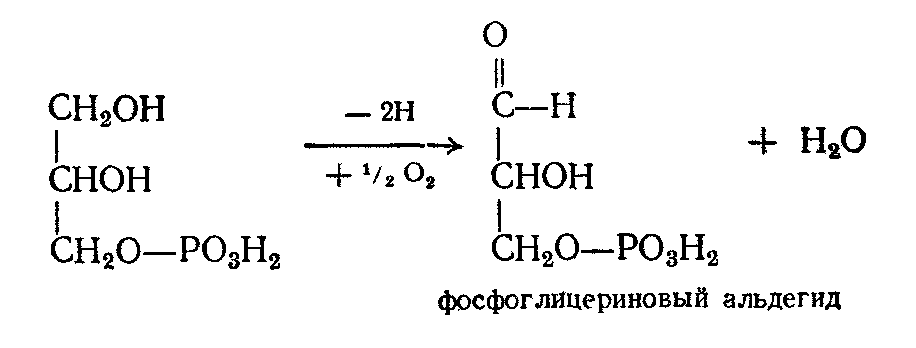
Фосфоглицериновый альдегид является одним из промежуточных продуктов гликолиза (см. превращения углеводов). Поэтому дальнейшие превращения протекают по схеме гликолиза, т.е. до лактата и пирувата, который превращается в ацетилКоА. Последний вступает в цикл трикарбоновых кислот, где ацетильный остаток окисляется до СО2 и Н2О.
Превращение жирных кислот.
В
основе современных представлений о
механизме окисления жирных кислот лежит
теория, предложенная Ф.Кноопом в 1904 г.
Им было экспериментально доказано, что
молекулы жирной кислоты распадаются
постепенно, укорачиваясь на 2 углеродных
атома. Эта теория получила название
теории
![]() -окисления,
потому что окислению всегда подвергается
второй от карбоксила углеродный атом
(находящийся, следовательно, в
-положении.
-окисления,
потому что окислению всегда подвергается
второй от карбоксила углеродный атом
(находящийся, следовательно, в
-положении.
В настоящее время установлено, что окислению жирных кислот в тканях предшествует их активация — взаимодействие с коферментом А в присутствии АТФ. Следствием этого является образование ацильного производного жирной кислоты, обладающего макроэргической связью:
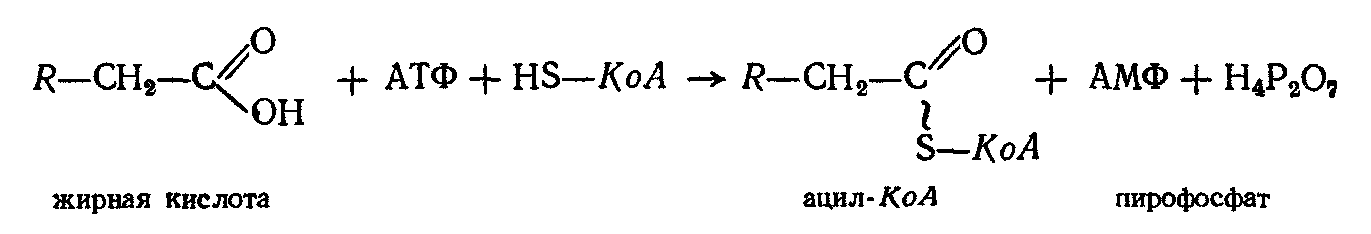
Активированная жирная кислота (ацил-КоА) дегидрогенируется, вследствие чего возникает двойная связь между атомами углерода. Реакция катализируется ферментами, представляющими собою флавопротеиды (ФлП), содержащие в качестве простетической группы ФАД.
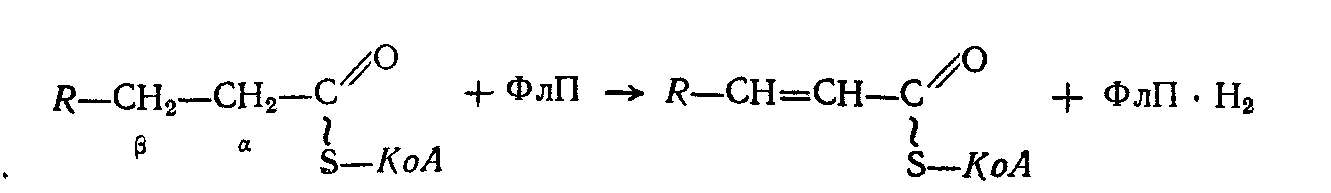
Затем по месту образования двойной связи, под влиянием фермента эноил-гидратазы, происходит присоединение молекулы воды (гидратация):
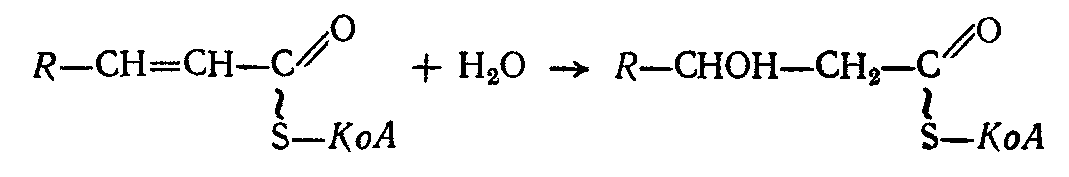
Образующий -гидроксиацилКоА подвергается второму дегидрогенированию, осуществляемому ферментом р-оксиацилдегидрогеназой, причем акцептирование водорода осуществляется его коферментом — окисленным НАД:
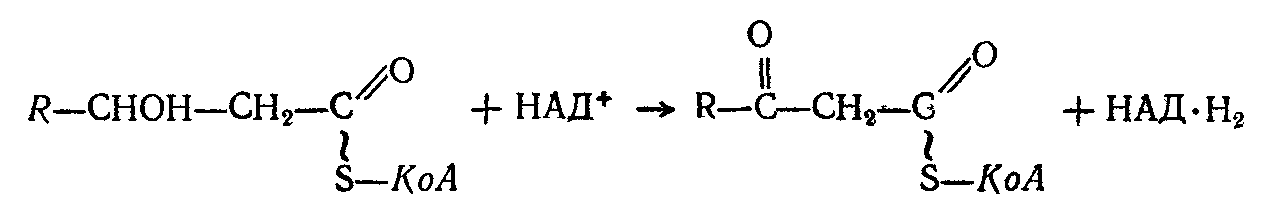
Продукт этой реакции — кетоацилКоА расщепляется с одновременным присоединением новой молекулы КоА к укороченной на 2 атома углерода жирной кислоте:
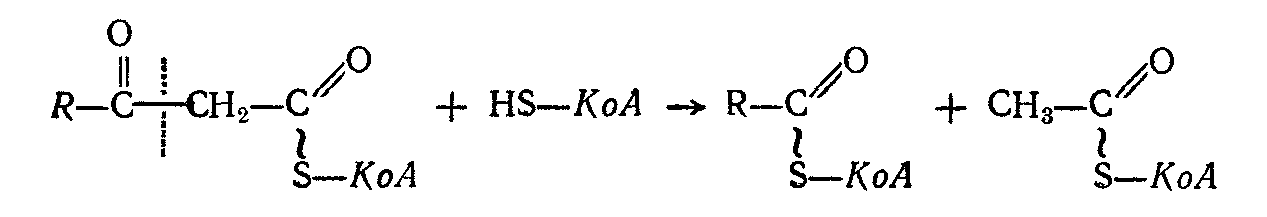
В результате этой реакции образуются ацетилКоА и новый жирный ацилКоА, содержащий на два углеродных атома меньше, чем исходная жирная кислота. Он, в свою очередь, может подвергаться описанным выше превращениям, так что в конечном итоге вся углеродная цепь жирной кислоты распадается на двууглеродные фрагменты ацетилКоА.
Образовавшийся при окислении жирных кислот ацетилКоА включается в цикл трикарбоновых кислот, где остаток уксусной кислоты окисляется до С02 и Н20, а КоА, освобождаясь, может вступать в конденсацию с новой молекулой жирной кислоты. При оксилении 1 молекулы ацетилКоА образуется 12 молекул АТФ. Кроме того, восстановленные ФАД и НАД, акцептирующие водород при дегидрогенированиях жирных кислот, окисляются в дыхательной цепи. При этом соответственно образуются 2 и 3 молекулы АТФ. В сумме полное окисление 1 двухуглеродного фрагмента любой жирной кислоты дает 17 молекул АТФ. Следовательно, при полном окислении 1 молекулы высокомолекулярной жирной кислоты, например пальмитиновой, образуется (8x17) 136 молекул АТФ. Из них 1 молекула расходуется на активацию жирной кислоты в самом начале процесса окисления.