

1) Трансформация нейрогуморальной сигнализации о метаболической потребности в процесс избирательного возбуждения структур мозга;
2) Процессы активации структур мозга как основы формирования соответствующего поведения.
Мотивационное возбуждение, возникшее первично в структурах гипоталамуса, распространяется до коры большого мозга, где происходит трансформация мотивационного возбуждения, вызванного метаболическими потребностями, в механизм целенаправленного поведения.
Доминирующая мотивация существенно изменяет свойства нейронов коры головного мозга. При этом повышается их функциональная мобильность, изменяется чувствительность к нейромедиаторам и нейропептидам.
Доминирующая мотивация также значительно повышает чувствительность соответствующих периферических рецепторов. При мотивации голода, например, возрастает чувствительность вкусовых рецепторов ротовой полости, при агрессивных мотивациях — рецепторов вокруг ротовой полости в тригеминальной области, при половых мотивациях избирательно активируются рецепторы половых органов. Все это расширяет взаимодействие субъектов, испытывающих ту или иную метаболическую потребность, с внешним миром и способствует более успешному удовлетворению исходных потребностей.
МОТИВАЦИЯ И ПАМЯТЬ
Мотивации тесно взаимодействуют с механизмами памяти. Мотивации, формирующиеся под воздействием внутренних метаболических потребностей и факторов окружаюшей среды, обладают выраженной способностью по опережающему принципу извлекать из памяти генетический и индивидуальный опыт субъектов по удовлетворению лежащей в их основе доминирующей потребности.
Восходящие активирующие влияния, сформированные различными мотивациями, запускают репликацию (удвоение) ДНК. Это приводит к синтезу специфического белка. А специфическтй белок участвует в формировании специфического поведения.
Говоря простым языком, повторные подкрепления приводят к тому что и процессы синтеза белка повторяются. В результате организм уже знает, какие ферменты ему необходимо «держать наготове» для синтеза того или иного специфического белка, который он уже производил не один раз. В результате получается, что подкрепления «тренируют» нейрональные ферменты синтеза специфического белка, что способствует закреплению памятного следа и опыт уже на молекулярном уровне, который возможно перевести затем в генетические формы памяти (врожжденные).
мотивация утрачивает способность трансформироваться в поведение при введении блокаторов синтеза белка вследствие подавления этого Синтеза.
Так, блокатор синтеза белка циклогексимид на несколько десятков часов подавляет пищевую мотивацию у кроликов при электрическом раздражении центра голода латерального гипоталамуса. После введения циклогексимида сначала облегчается, а через несколько часов подавляется оборонительная реакция кроликов при электрическом раздражении центра страха вентро-медиального гипоталамуса.
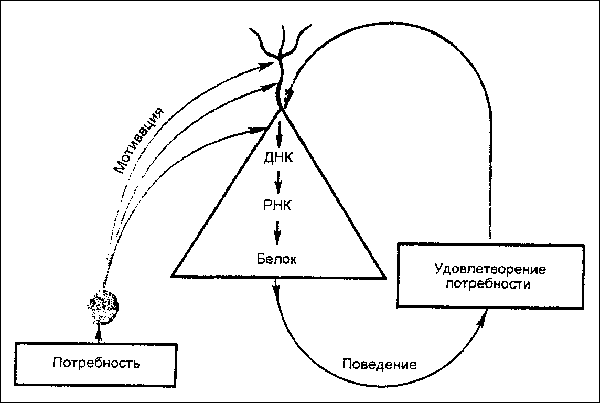
Рис.2 Молекулярные изменения в генетическом аппарате нейрона коры под влиянием восходящих активируюших мотивационных возбуждений. Повторное удовлетворение потребности инициирует синтез в теле нейрона и белка.
На фоне действия циклогексимида пентагастрин, введенный в боковые желудочки мозга, восстанавливает пищевую мотивацию. Пищевую реакцию у кроликов при электрическом раздражении латерального гипоталамуса блокируют также иммуноглобулины к гастрину, связывающие гастрин в нерастворимый комплекс.
Имеется множество физиологических экспериментов, которые показывают, что доминирующая мотивация извлекает у животных опыт, ранее накопленный по удовлетворению соответствующих потребностей, по опережающему принципу до конечного подкрепляющего результата включительно и, таким образом, как бы "вытягивает" весь предшествующий опыт животного по удовлетворению соответствующей потребности.
В опытах кроликов предварительно перед получением пищи обучали совершать специальные ритуальные движения — поворот на 360°. В процессе выработки пищедобывателъных навыков каждый такой поворот подкреплялся порцией пищи. В результате уже к 30—40-му сочетанию животные демонстрировали прочно выработанный пишедобывающий навык: при помещении в экспериментальную камеру они совершали повороты на 360°, которые во всех случаях подкреплялись пищей. После того как у всех животных был упрочен пищедобывательный навык предварительного поворота, каждому из них в область латерального гипоталамуса вживляли биполярные электроды. Кончик электрода фиксировали в той точке латерального гипоталамуса, раздражение которой у накормленных животных вызывало отчетливо выраженную реакцию дополнительного приема находящейся перед ними пищи.
Вопрос заключается в следуюшем: способно ли искусственное раздражение мотивациогенного центра голода латерального гипоталамуса выявить не только пищевую реакцию приема пищи, но и то ритуальное движение на 360°, которому каждое из подопытных животных было обучено в предварительных экспериментах?
Опыты были проведены на кроликах в условиях той же экспериментальной камеры, в которой они предварительно обучались ритуальным пищедобывательным движениям.
Эксперименты показали следующее. Все животные, накормленные перед опытом, в экспериментальной клетке демонстрировали полное успокоение. Однако вслед за стимуляцией латерального гипоталамуса они оживлялись и совершали ритуальное движение — поворот на 360°, после чего устремлялись к кормушке и поедали находящуюся в ней порцию пищи.
Опыты свидетельствуют о том, что доминирующая пищевая мотивация, вызванная у накормленных животных искусственным раздражением латерального гипоталамуса, привела к мобилизации всего того опыта, который в данной обстановке в предшествующем обучении предварял пищевое подкрепление. Это положение подтверждается опытом противоположного смысла. У животных другой группы с выработанным аналогичным ритуальным движением производили двустороннюю коагуляцию пищевых центров латерального гипоталамуса. Оказалось, что такая операция полностью элиминировала в экспериментальной камере как ритуальное движение, так и прием пиши. Опыты в этой серии с очевидностью указывают на ведущее значение доминирующей мотивации в извлечении навыка из памяти, причем извлечение опыта из памяти происходит до конечного результата включительно.
Кроме извлечения опыта из памяти, доминируюшая мотивация определяет и быстроту фиксации промежуточных и конечных результатов действия по удовлетворению соответствующих потребностей (пример с тренировкой в настоящем спарринге или в «бое с тенью»).
В экспериментах Т. Н. Ониани с сотр. был проведен следуюший опыт. Голодных кошек помешали в специальную камеру, разделенную на два отсека. В первом отсеке животные не получали пищу, а должны были зафиксировать местоположение светового сигнала справа или слева, что соответствовало расположению кормушек с пищей во втором отсеке камеры. Вход во второй отсек преграждала дверь. Опыт проходил таким образом, что вслед за подачей правого или левого светового сигнала через определенный промежуток времени открывалась дверца во второй отсек и животноё получало пищу в кормушке, находяшейся на стороне сигнала. Путем тренировки у животных вырабатывали четкую линию поведения. При включении правой лампочки кошки при открывании двери во второй отсек устремлялись к правой кормушке, при зажигании левой лампочки — к левой кормушке. После этого замысел эксперимента усложнялся. Между подачей светового сигнала и открытием дверцы изменяли интервалы времени. Определялось максимальное время отсроченных реакций, при котором животные удерживали след сигнала и не совершали ошибок в выборе стороны расположения кормушки во втором отсеке. Оказалось, что в голодном состоянии при наличии выраженной пищевой мотивации время отсроченных реакций могло быть значительным. Однако оно существенно сокращалось по мере насыщения животных, удовлетворения их ведущей потребности и снижения доминирующей мотивации. Эксперимент отчетливо демонстрирует значение мотивацил для удержания следов памяти.
Итак, в системной организации целенаправленных поведенческих актов мотивации выступают как ведущий фактор фиксации опыта в памяти, способности его удержания при наличии препятствий к достижению цели и, наконец, обусловливают быстрое извлечение накопленного опыта из памяти.
Из этого следует важная практическая рекомендация: запоминание всегда успешно при наличии выраженной мотивации. Лучше запоминается тот предмет, который представляет для обучающегося наиболее значимый интерес.
С физиологической точки зрения, процессы фиксации доминирующей мотивации опыта по удовлетворению соответствующей потребности изучены недостаточно.
НАПРАВЛЯЮЩИЙ КОМПОНЕНТ ДОМИНИРУЮЩЕЙ МОТИВАЦИИ
Практически все основные биологические мотивации (пищевые, питьевые, половые, оборонительные, агрессивные etc.) генетически программируют ведущие свойства соответствующих подкрепляющих раздражителей, которые становятся УЗНАВАЕМЫМИ ПРИ ПРЕДЪЯВЛЕНИИ. Только благодаря этому новорожденные животные проявляют столь поразительное избирательное отношение к биологически значимым раздражителям окружаюшей среды, которое затем совершенствуется в процессах индивидуального обучения. Опережающее программирование свойств подкрепляющих раздражителей осуществляется в аппарате акцептора результата действия.
С этим формируюшимся по опережающему принципу направляюшим компонентом доминирующей мотивации постоянно происходит сравнение параметров реально достигнутых результатов. Этот механизм в конечном счете и направляет животных к полноценному удовлетворению их ведущих потребностей и позволяет им оценивать и исправлять ошибки поведенческой деятельности.
МОТИВАЦИИ И ПОДКРЕПЛЕНИЕ
В системной организации поведения доминирующая мотивация и подкрепление тесно взаимодействуют, причем часто на одних и тех же нейронах мозга. Подкрепляющее возбуждение изменяет активность нейронов, вовлеченных в исходную мотивацию. При этом пачкообразная активность этих нейронов сменяется упорядоченной, регулярной (см. рис.).
Реакции других нейронов мозга, не вовлеченных в доминирующую мотивацию, на подкрепляющее возбуждение могут быть самыми разнообразными — в виде учащения или торможения исходной активности, но не упорядоченно.
Следовательно, мотивационное доминирующее возбуждение как бы "настраивает" нейроны различных областей мозга на подкрепляющее воздействие. В то же время подкрепление изменяет также реакции нейронов мозга на мотивационные воздействия.
Взаимодействие мотивационных и подкрепляющих возбуждений на отдельных нейронах мозга строится по комплементарному принципу (поэтому при пищевом подкреплении рефлекса животное должно быть несколько голодным). На них существенное влияние оказывают иммуномодуляторы и нейропептиды.
Доминирующее мотивационное возбуждение, представляет собой функциональную канву избирательно возбужденных синаптических и нейрональных образований мозга.
На этой канве подкрепляющие этапные и завершающие возбуждения в определенной временной последовательности пишут своеобразный узор, или энграмму. Этот узор по мере неоднократных подкреплений отшлифовывается в форме специфической корково-подкорковой архитектуры, в которой синаптические и нейрональные элементы объединены в пространственно-временных соотношениях. Система запоминает эту архитектуру чтобы еще раз не строить ее аново. Для экономии времени.
При каждом очередном возникновении соответствующей потребности доминируюшее мотивационное состояние активирует элементы выработанной на основе предшествующего опыта энграммы (оно же проходит по этим путям, поэтому активация неизбедна), возбуждая их до конечного пункта, связанного с получением необходимой информации об удовлетворении соответствующей потребности. Этот комплекс избирательно возбужденных корково-подкорковых аппаратов, представляющий нейрофизиологическую архитектуру акцептора результатов действия, и направляет поведение живого существа через постоянное сравнение поступающей к нему с периферии обратной афферентации, вызванной действием раздражителей внешней среды, к достижению цели, т. е. к удовлетворению доминирующей на каждый данный момент времени потребности.
БИОЛОГИЧЕСКИЕ МОТИВАЦИИ В ФОРМИРОВАНИИ ЛИЧНОСТИ
Выраженная мотивация человека обусловливает целеустремленность личности, ее способность активно действовать для достижения целей в соответствии с юридическими, правовыми и моральными законодательствами общества.
Врожденные биологические мотивации также участвуют в формировании личности, определяя ее индивидуальные и общественные интересы, а также черты характера уже в раннем детстве.
По доминированию пищевых, агрессивных, оборонителъных, половых и других биологических мотиваций выявляются индивидуальные характеры.
Тем не менее, несмотря на важное значение биологических мотиваций, ведущее значение в формировании личности человека, его характерологических особенностей принадлежит социальным мотивациям, формирующимся под влиянием окружающей, в частности социальной, среды. Вопросы формирования личности человека широко освещаются в соответствуюшей психологической литературе.
ПАТОЛОГИЧЕСКИЕ МОТИВАЦИИ
К патологическим относятся такие искусственно создаваемые влечения, как наркомании, алкоголизм и курение. Имеются и другие патологические влечения, которые рассматриваются в курсе психиатрии.
В случае употребления алкоголя и наркотиков в гипоталамических структурах мозга на основе изменения метаболических реакций формируются искусственные пейсмекеры, создаюшие в отсутствие наркотиков активное возбуждение структур мозга, приводящее к выраженному влечению к их употреблению (см. рис.).

Рис. Динамика формирования алкогольной мотивации на основе биологической мотивации жажды. а — осмотическая потребность приводит к формированию мотивации жажды, к поиску и приему воды; б — под влиянием приема этанола изменяются свойства "центров жажды" гипоталамуса: они прекращают оказывать активирующие влияния на кору; в — осмотическая потребность формирует патологическую алкогольную мотивацию.
Расстройства мотиваций
Расстройства мотиваций проявляются в их усилении (булимия, гиперсексуальность, полидипсия) или значительном подавлении (афагия, адипсия, импотенция). В случаях избирательного расстройства какой-либо одной, преимущественно врожденной, мотивации нередко происходит формирование психопатической личности.
Представление о формировании мотиваций по пейсмекерному принципу определяет возможность избирательно направленного воздействия на патологические их формы. Фармакологические и другие виды воздействий должны быть направлены прежде всего на метаболизм пейсмекеров, имеющих, как указывалось выше, повышенную чувствительность по сравнению с другими элементами мотивационного возбуждения. Именно путем таких направленных воздействий на самое уязвимое звено патологической мотивации можно разрушить всю систему мотивационного возбуждения, включая связанные с ней формы патологических поведенческих реакций и патологические навыки.
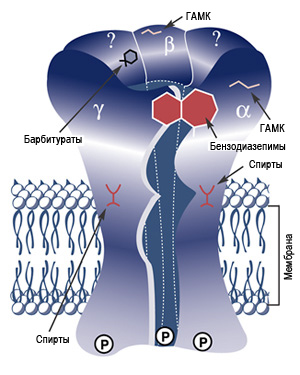
На рисунке изображено схематически строение ГАМК-бензодиазепино-барбитурового рецептурного комплекса.
Значение лимбической системы в формировании мотиваций
Она подключена к разным мотивационным механизмам и обеспечивает оценку поведения на основе принципа «лучше — хуже» и направление поведения в какую либо сторону.
Экспериментально доказано, что эмоциональное поведение животного при стимуляции некоторых участков лимбической системы проявляется главным образом реакциями агрессии (гнева), убегания (страха) или наблюдаются смешанные формы поведения, например оборонительные реакции.
Эмоции в отличие от мотиваций возникают в ответ на внезапные изменения среды , скоротечны и факультативны. Длительные немотивированные изменения эмоционального поведения могут быть следствием органической патологии или действия некоторых нейролептиков.
В разных отделах лимбической системы открыты центры «удовольствия» и «неудовольствия», объединенные в системы «награды» и «наказания». При стимуляции системы «наказания» животные ведут себя так же, как при страхе или боли, а при стимуляции системы «награда» стремятся возобновить раздражение и осуществляют его самостоятельно, если им представляется такая возможность. Т.е. при возникновении эмоций
Эффекты награды непосредственно не связаны с регуляцией биологических мотиваций или торможением отрицательных эмоций и скорее всего представляют неспецифический механизм положительного подкрепления, деятельность которого воспринимается как удовольствие или награда.
Висцеральные реакции при воздействии на лимбическую систему, как правило, являются специфическим компонентом соответствующего типа поведения. Так, при стимуляции центра голода в латеральных отделах гипоталамуса наблюдаются обильное слюноотделение, усиление моторики и секреторной активности желудочно-кишечного тракта; при провокации половых реакций — эрекция, эякуляция и т.д., а в целом на фоне разных типов мотивационного и эмоционального поведения регистрируются изменения дыхания, частоты сердечных сокращений и величины АД, секреции АКТГ, катехоламинов, других гормонов и медиаторов,
Для объяснения принципов интегративной деятельности лимбической системы выдвинуто представление о циклическом характере движения процессов возбуждения по замкнутой сети структур, включающих гиппокамп, сосцевидные тела, свод мозга, передние ядра таламуса, поясную извилину — так называемый круг Пейпса. Далее цикл возобновляется. Этот «транзитный» принцип организации функций лимбической системы подтверждается рядом фактов. Например, пищевые реакции удается вызвать при стимуляции латерального ядра гипоталамуса, латеральной преоптической области и некоторых других структур. Тем не менее несмотря на множественность локализации функций удалось установить ключевые, или пейсмекерные, механизмы, выключение которых ведет к полному выпадению функции.
В настоящее время проблема консолидации структур в определенную функциональную систему решается с позиций нейрохимии. Показано, что многие образования лимбической системы содержат клетки и терминали, секретирующие несколько типов биологически активных веществ. Среди них наиболее изучены моноаминергические нейроны, образующие три системы: дофаминергическую, норадренергическую и серотонинергическую. Нейрохимическое сродство отдельным структур лимбической системы во многом предопределяет степень их участия в том или ином типе поведения. Деятельность системы награды обеспечивается норадренергическими и дофаминергическими механизмами; блокада соответствующих клеточных рецепторов препаратами из ряда фенотиазинов или бугарофенонов сопровождается эмоциональной и двигательной заторможенностью, а при избыточных дозировках — депрессией и двигательными нарушениями, близкими к синдрому паркинсонизма. В регуляции сна и бодрствования, наряду с моноаминергическими механизмами, участвуют ГАМК-эргические и нейромодуляторные механизмы, специфически реагирующие на гамма-аминомасляную кислоту (ГАМК) и пептид дельта-сна. В механизмах боли ключевую роль играют эндогенная опиатная система и морфиноподобные вещества — эндорфины и энкефалины.
Нарушения функций лимбической системы проявляются при разных заболеваниях (травмах мозга, интоксикациях, нейроинфекциях, сосудистой патологии, эндогенных психозах, неврозах) и бывают чрезвычайно разнообразными по клинической картине. В зависимости от локализации и объема поражения эти расстройства могут иметь отношение к мотивациям, эмоциям, вегетативным функциям и сочетаться в разных пропорциях. Низкие пороги судорожной активности лимбической системы обусловливают разные формы эпилепсии: большие и малые формы судорожных припадков, автоматизмы, изменения сознания (деперсонализация и дереализация), вегетативные пароксизмы, которым предшествуют или сопутствуют разные формы изменения настроения в сочетании с обонятельными, вкусовыми и слуховыми галлюцинациями.
3. Роль лимбической системы в формировании мотиваций, эмоций, организации памяти
Взаимное подавление этих частей мозга происходит по многим направлениям. Лимбическая система и новые области коры головного мозга подавляют Р-комплекс, но под влиянием жизни в обществе может случиться также, что Р-комплекс станет угнетать новые области коры, а одно полушарие главенствовать над другим.
Английский антрополог Бернард Кемпбелл из Кембриджского университета предполагает, что лимбическая система достаточно хорошо связана с правым полушарием коры головного мозга, которое, как мы видели, намного лучше, чем левое, управляет эмоциями.. Болезнь Ги де ля Туретта, при всей своей сложности - скорее всего есть результат недостатка некоторого химического вещества, передающего информацию от нейрона к нейрону, и тщательно подобранные дозы галоперидола сильно облегчают вызываемые ею страдания.
Последние данные указывают, что такие лимбические гормоны, как АКТГ (адренокортикотропный гормон) и вазопрессин, могут сильно улучшить способность животных удерживать и вызывать в памяти различные факты. Этот и аналогичные примеры указывают путь если не к решительному со- вершенствованию мозга, то хотя бы к его существенному улучшению - быть может, с помощью уменьшения избытка или регулирования производства в мозге пептидных молекул определенных типов.
Подобные примеры также в высшей мере снижают бремя вины, которое, как правило, испытывают те, кто страдает умственными расстройствами,- бремя, крайне редко знакомое, скажем, больным корью.
Уже сейчас есть ряд психотропных и влияющих на настроение средств, которые в различной степени опасны для человека (этиловый спирт среди них - одно из наиболее вредных и широко распространенных) и которые воздействуют на специфические части Р-комплекса, лимбической системы и неокортекса. Есть основания полагать, что многие алкалоиды и другие средства, влияющие на человеческое поведение, оказывают свое действие потому, что они химически близки к некоторым небольшим естественным пептидным молекулам мозга, например эндорфинам. Многие из этих пептидов воздействуют на лимбическую систему и связаны с нашим эмоциональным состоянием.
Мозг человека отражает реальные параметры подкрепляющих воздействий на организм на информационной основе. Поступающие в мозг от многочисленных периферических рецепторов сигналы многократно преобразуются - сначала в форму нервных импульсаций проводящих нервных волокон и в определенный набор информационных гуморальных молекул, затем в специфическое возбуждение определенных структур мозга. Однако информационный смысл разных свойств подкрепления, несмотря на многочисленную смену форм физико-химических и физиологических процессов, остается неискаженным. С другой стороны, генетический и индивидуально приобретенный опыт субъектов по удовлетворению их различных потребностей определяется образованными информационными молекулами - молекулярными энграммами акцепторов результатов действия разных функциональных систем, осуществляющих оценку поступающей к ним информации и участвующих в организации соответствующих психических и поведенческих актов.
Каждый параметр подкрепляющего воздействия на лимбическую систему оставляет свой специфический информационный след в соответствующей зрительной, вкусовой, слуховой, тактильной и любой другой проекционной зоне мозга, определяя тем самым генерализованную по различным структурам мозга архитектонику акцептора результата действия.
При этом если генетические компоненты акцепторов результатов действия разных функциональных систем консервативны, то в процессе индивидуальной жизни их архитектоника все время динамически изменяется в соответствии с изменчивостью параметров подкрепляющих воздействий. Каждое подкрепление как часть многогранной действительности оставляет свой специфический информационный след на структурах акцептора результата действия лимбической системы.
При стабильных условиях существования и однотипных формах подкрепления <отпечатки действительности> на структурах соответствующих акцепторов результатов действия формируют устойчивые стереотипы, которые могут сохраняться длительное время, а иногда и всю жизнь индивида. В изменяющихся условиях жизни <отпечатки действительности> неустойчивы и быстро сменяют друг друга. <Отпечатки действительности> на структурах акцепторов результатов действия разных функциональных систем направляют поведение человека и животных на оптимальное удовлетворение их насущных потребностей, выступая в роли своеобразных векторов поведения.
Физико-химические процессы, разыгрывающиеся в нейронах акцептора результата действия под влиянием доминирующей мотивации, порождают информационный процесс опережающего возбуждения - предвидения свойств потребных результатов и способов их достижения. Таким образом, материальная потребность трансформируется в идеальный информационный процесс.
В свою очередь возникающие на основе доминирующей мотивации поведение и взаимодействие субъектов с объектами внешнего мира определяют трансформацию информационных процессов в материальные. Оценка параметров достигнутых результатов снова осуществляется на информационной основе обратной афферентации и сравнения ее с акцептором результата действия. Следовательно, мозг, постоянно взаимодействующий с действительностью, осуществляет непрерывную трансформацию информационных процессов без потери их информационной значимости.
Мозг человека и животных постоянно строит <отпечатки действительности>. Они включают образы подкрепляющих воздействий и соответствующие программы поведения. Все эти процессы на информационной основе с помощью генетических и приобретенных механизмов памяти происходят на структурах акцептора результата действия.
При этом мозг животных строит <отпечатки действительности>, по-видимому, только на эмоциональной основе, в то время как человек приобрел в процессе эволюции качественно новую - языковую форму построения <отпечатков действительности>.
Особенностью лимбической системы является то, что между ее структурами имеются простые двусторонние связи и сложные пути, образующие множество замкнутых кругов (см. рис. 4.12). Такая организация создает условия для длительного циркулирования одного и того же возбуждения в системе и тем самым для сохранения в ней единого состояния и навязывание этого состояния другим системам мозга.
В настоящее время хорошо известны связи между структурами мозга, организующие круги, имеющие свою функциональную специфику. К ним относится круг Пейпеса (гиппокамп à сосцевидные тела à передние ядра таламуса à кора поясной извилины à парагиппокампова извилина à гиппокамп). Этот круг имеет отношение к памяти и процессам обучения.
Другой круг (миндалевидное тело à гипоталамус à мезенцефальные структуры à миндалевидное тело) регулирует агрессивно-оборонительные, пищевые и сексуальные формы поведения.
Считается, что образная (иконическая) память формируется кортико-лимбико-таламо-кортикальным кругом. Круги разного функционального назначения связывают лимбическую систему со многими структурами центральной нервной системы, что позволяет последней реализовать функции, специфика которых определяется включенной дополнительной структурой.
Например, включение хвостатого ядра в один из кругов лимбической системы определяет ее участие в организации тормозных процессов высшей нервной деятельности.
Большое количество связей в лимбической системе, своеобразное круговое взаимодействие ее структур создают благоприятные условия для реверберации возбуждения по коротким и длинным кругам. Это, с одной стороны, обеспечивает функциональное взаимодействие частей лимбической системы, с другой — создает условия для запоминания. Обилие связей лимбической системы со структурами центральной нервной системы затрудняет выделение функций мозга, в которых она не принимала бы участия. Так, лимбическая система имеет отношение к регулированию уровня реакции автономной, соматической систем при эмоционально-мотивационной деятельности, регулированию уровня внимания, восприятия, воспроизведения эмоционально значимой информации. Лимбическая система определяет выбор и реализацию адаптационных форм поведения, динамику врожденных форм поведения, поддержание гомеостаза, генеративных процессов. Наконец, она обеспечивает создание эмоционального фона, формирование и реализацию процессов высшей нервной деятельности.
Нужно отметить, что древняя и старая кора лимбической системы имеет прямое отношение к обонятельной функции. В свою очередь обонятельный анализатор, как самый древний из анализаторов, является неспецифическим активатором всех видов деятельности коры большого мозга.
Некоторые авторы называют лимбическую систему висцеральным мозгом, т. е. структурой ЦНС, участвующей в регуляции деятельности внутренних органов. И действительно, миндалевидные тела, прозрачная перегородка, обонятельный мозг при их возбуждении изменяют активность вегетативных систем организма в соответствии с условиями окружающей среды. Это стало возможно благодаря установлению морфологических и функциональных связей с более молодыми образованиями мозга, обеспечивающими взаимодействие экстероцептивных, интероцептивных систем и коры височной доли.
Наиболее полифункциональными образованиями лимбической системы являются гиппокамп и миндалевидные тела. Физиология этих структур наиболее изучена
Влияние двигательной активности на регуляторные механизмы ЦНС и гормонального звена
И. П. Павлов отмечал, что двигательная деятельность приносит человеку «мышечную радость». Во время движений ЦНС перерабатывает большой объем информации, связанный с проприоцептивной афферентацией от мышц. Функциональное состояние всех отделов ЦНС, в том числе коры больших полушарий и подкорковых центров, повышается. Активизируются и балансируются возбудительный и тормозной процессы. Укорачивается время двигательной реакции на звуковые и световые раздражители, повышается частота усвоения ритма раздражений, усиливается выраженность альфа-ритма в состоянии покоя. В клетках коры больших полушарий увеличивается содержание РНК, имеющей непосредственное отношение к механизмам памяти. Усиливаются ассоциативные процессы, возникают «озарения», составляющие физиологическую основу экстраполяции (творчества). За счет корковых посылок и рефлекторных влияний с периферии активизируется деятельность гипоталамо-гипофизарной системы. При этом в реакцию вовлекается эндокринная система и достигается оптимальная (соответственно уровню двигательной активности) регуляция гемодинамики, дыхания, кроветворения, выделительной функции почек, кишечника, дезинтоксикационной функции печени. Во время бега и других видов двигательной активности в кровь выделяются гормоны, эндорфины. Они уменьшают уровень тревожности, подавляют чувство страха, боли и голода.
За счет увеличения функциональных резервов организма повышается его жизненный тонус. Возрастают устойчивость к стрессорным факторам, физическая и психическая работоспособность.
Влияние двигательной активности на функции нервно-мышечного аппарата
В зависимости от характера двигательной активности скелетные мышцы человека могут работать в динамическом, статическом и смешанном режимах. Во время движения в связи с повышением уровня обмена веществ в соответствии с правилом А. Крога в мышцах увеличивается число открытых капилляров. Увеличенный приток крови к мышцам способствует повышению их температуры, что обусловливает уменьшение вязкости (силы трения между отдельными волокнами), а следовательно, облегчает реализацию физико-химических свойств мышц, непосредственно влияющих на производительность совершаемой работы. При статических усилиях сосуды мышц сдавливаются находящимися в состоянии напряжения волокнами, кровообращение в мышце почти прекращается. То небольшое количество О2, которое находится в составе миоглобина, не может поддерживать аэробный режим энергообеспечения, в связи с чем преобладает анаэробный режим с использованием креатинфосфокиназной реакции и гликолитического фосфорилирования.
Систематическая двигательная деятельность вызывает рабочую гипертрофию мышечных волокон, увеличение емкости капиллярной сети в мышцах, содержания миоглобина, гликогена, АТФ, КФ, дыхательных ферментов. В волокнах повышается количество митохондрий. Последние способствуют возрастанию способности мышц утилизировать пируват. При этом ограничивается накопление молочной кислоты и обеспечивается возможность мобилизации жирных кислот, повышается способность к интенсивной и длительной мышечной работе. Параллельно наступают изменения в центральном звене двигательных единиц — в α-мотонейронах, которые гипертрофируются при одновременном увеличении содержания в них дыхательных ферментов. При статическом режиме деятельности мышц в них происходит более глубокая перестройка сосудистой системы и нервных окончаний: капилляры изменяют ход — идут не параллельно мышечным волокнам, а оплетают их, аксоны нейронов двигательных единиц делятся на большее количество терминален, подходящих к мышечным волокнам. Надежность функционирования опорно-двигательного аппарата возрастает за счет увеличения поперечника трубчатых костей и утолщения их компактного вещества.
По мере повторения моторных нагрузок двигательная функциональная система приобретает все большую надежность деятельности. Это выражается в совершенствовании координации, автоматизации и экономичности движений. В основе этого лежат расширение межцентральных связей различных моторных уровней коры больших полушарий, стриопаллидарной системы, среднего, продолговатого мозга, а также формирование динамического стереотипа с высокой помехоустойчивостью.
Научно обоснованная двигательная деятельность в виде занятий физической культурой способствует правильному формированию осанки, адекватному развитию мышечного «корсета» в период интенсивного роста, особенно в пубертатный период, характеризующийся ростовым скачком.
2,03 12 галетина
Головко
Коржов
Суюнова
Зленко
гулиева