

Модель а.Д.Базыкина
Теоретический анализ моделей взаимодействий видов наиболее исчерпывающе проведен в книге А.Д.Базыкина “Биофизика взаимодействующих популяций” (М., Наука, 1985).
Рассмотрим одну из изученных в этой книге моделей типа хищник-жертва.
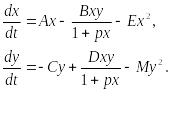 (9.17)
(9.17)
Система (9.17) является обобщением простейшей модели хищник-жертва Вольтерра (5.17) с учетом эффекта насыщения хищников. В модели (5.17) предполагается, что интенсивность выедания жертв линейно растет с ростом плотности жертв, что при больших плотностях жертв не соответствует реальности. Для описания зависимости рациона хищника от плотности жертв могут быть выбраны разные функции. Наиболее существенно, чтобы выбранная функция с ростом x асимптотически стремилась к постоянному значению. В модели (9.6) использовалась логистическая зависимость. В модели Базыкина в роли такой функции выбрана гипербола x/(1+px). Вспомним, что такой вид имеет формула Моно, описывающая зависимость скорость роста микроорганизмов от концентрации субстрата. Здесь в роли субстрата выступает жертва, а в роли микроорганизмов – хищник.
Система (9.17) зависит от семи параметров. Число параметров можно уменьшить с помощью замены переменных:
x®(A/D)x; y®(A/D)/y;
t®(1/A)t; g=c/A;
a=PD/A; e=E/D; m=M/B
В новых безразмерных переменных система имеет вид
 (9.18)
(9.18)
и зависит от четырех параметров.
Для полного качественного исследования необходимо разбить четырехмерное пространство параметров на области с различным типом динамического поведения, т.е. построить параметрический, или структурный портрет системы.
Затем надо построить фазовые портреты для каждой из областей параметрического портрета и описать бифуркации, происходящие с фазовыми портретами на границах различных областей параметрического портрета.
Построение полного параметрического портрета производится в виде набора “срезов” (проекций) параметрического портрета малой размерности при фиксированных значениях некоторых из параметров.
Параметрический портрет системы (9.18) при фиксированных g и малых e представлен на рис.9.8. Портрет содержит 10 областей с различным типом поведения фазовых траекторий.
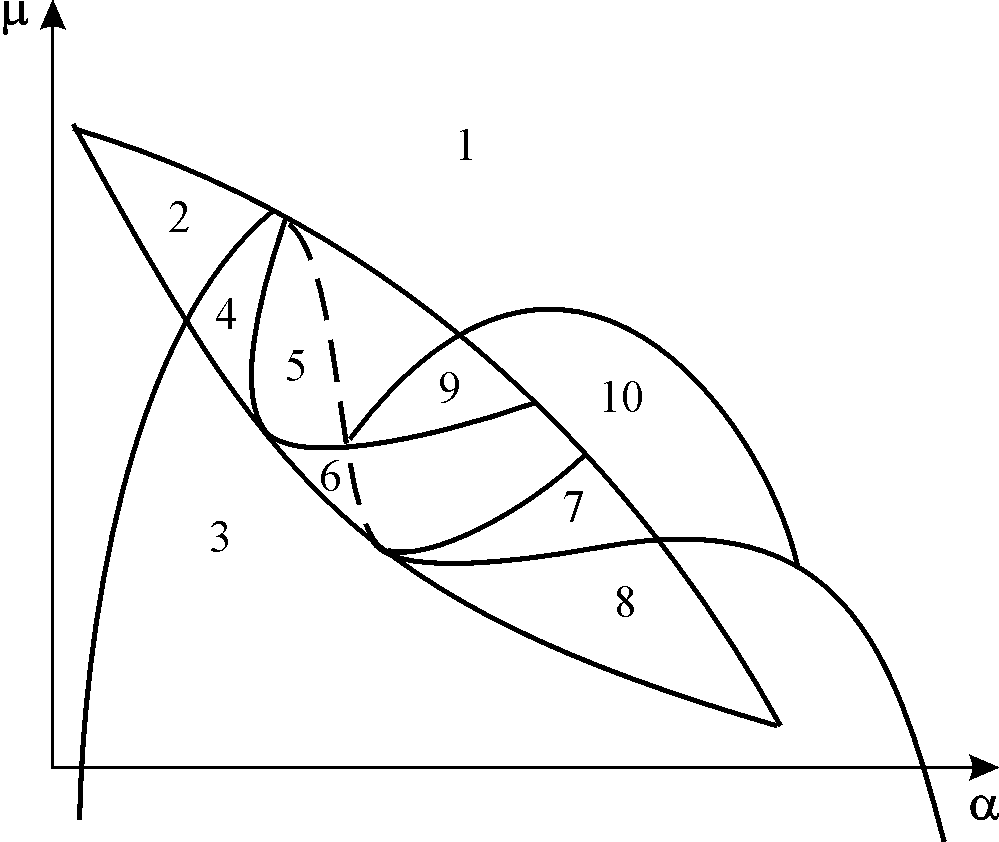
Рис. 9.8. Параметрический портрет системы (9.18) при фиксированных g
и малых e
Поведение системы при различных соотношениях параметров может быть существенно различным (рис.9.9). В системе возможны:
1) одно устойчивое равновесие (области 1 и 5);
2) один устойчивый предельный цикл (области 3 и 8);
3) два устойчивых равновесия (область 2)
4) устойчивый предельный цикл и неустойчивое равновесие внутри него (области 6, 7, 9, 10)
5) устойчивый предельный цикл и устойчивое равновесие вне его (область 4).
В параметрических областях 7, 9, 10 область притяжения равновесия ограничивается неустойчивым предельным циклом, лежащим внутри устойчивого. Наиболее интересно устроен фазовый портрет, соответствующий области 6 на параметрическом портрете. Детально он изображен на рис. 9.10.
Рис. 9.9. Набор фазовых портретов системы (9.18), возможных в конечной части первого квадранта и соответствующих областям 1 ‑ 10 параметрического портрета рис. 9.8 (Базыкин, 1985)
|
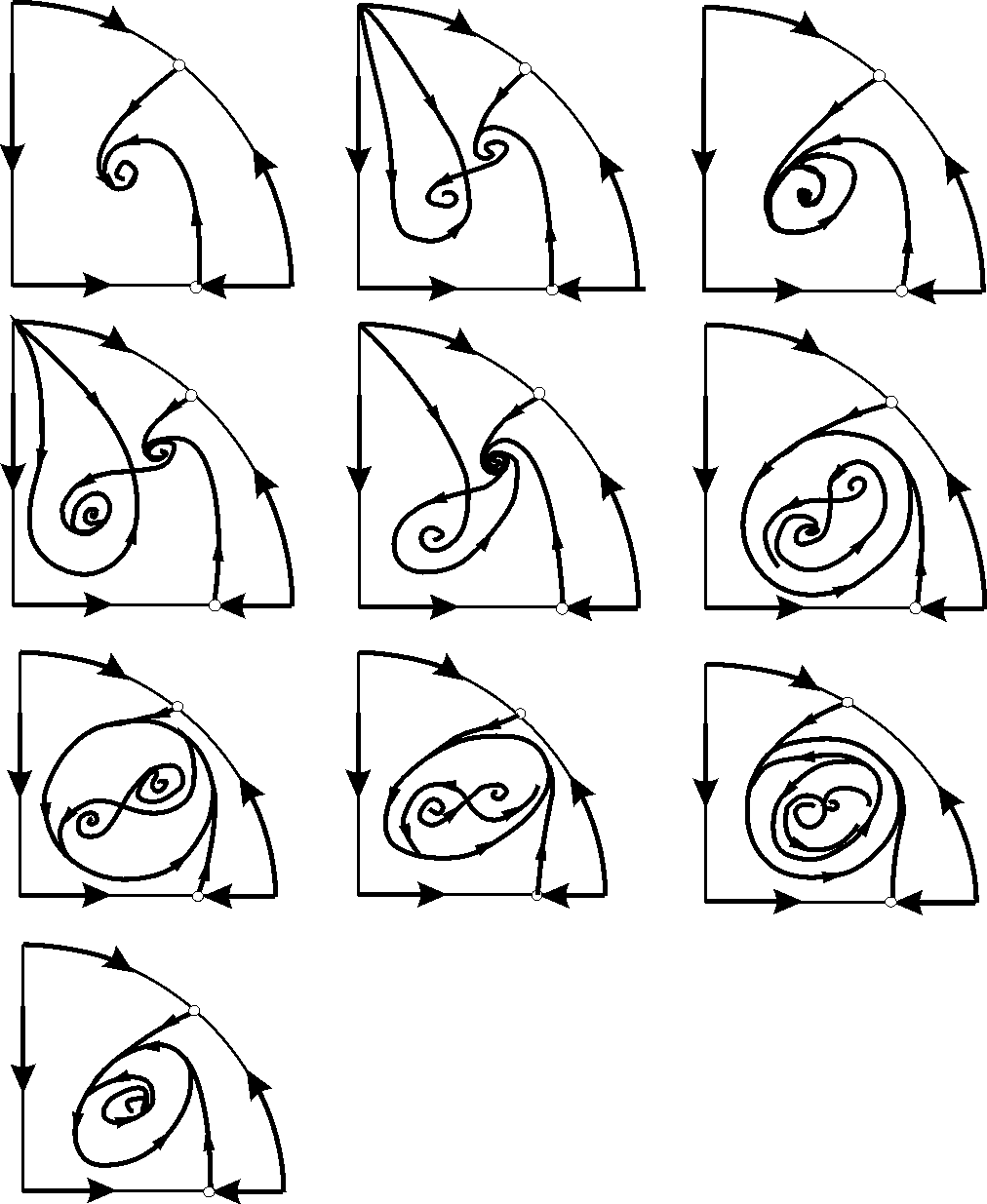
Область притяжения равновесия В2 (заштрихована) представляет собой “улитку”, скручивающуюся с неустойчивого фокуса В1. Если известно, что в начальный момент времени система находилась в окрестности В1, то судить о том, придет ли соответствующая траектория в равновесие В2 или на устойчивый предельный цикл, окружающий три точки равновесия С (седло), В1 и В2 можно лишь на основе вероятностных соображений.
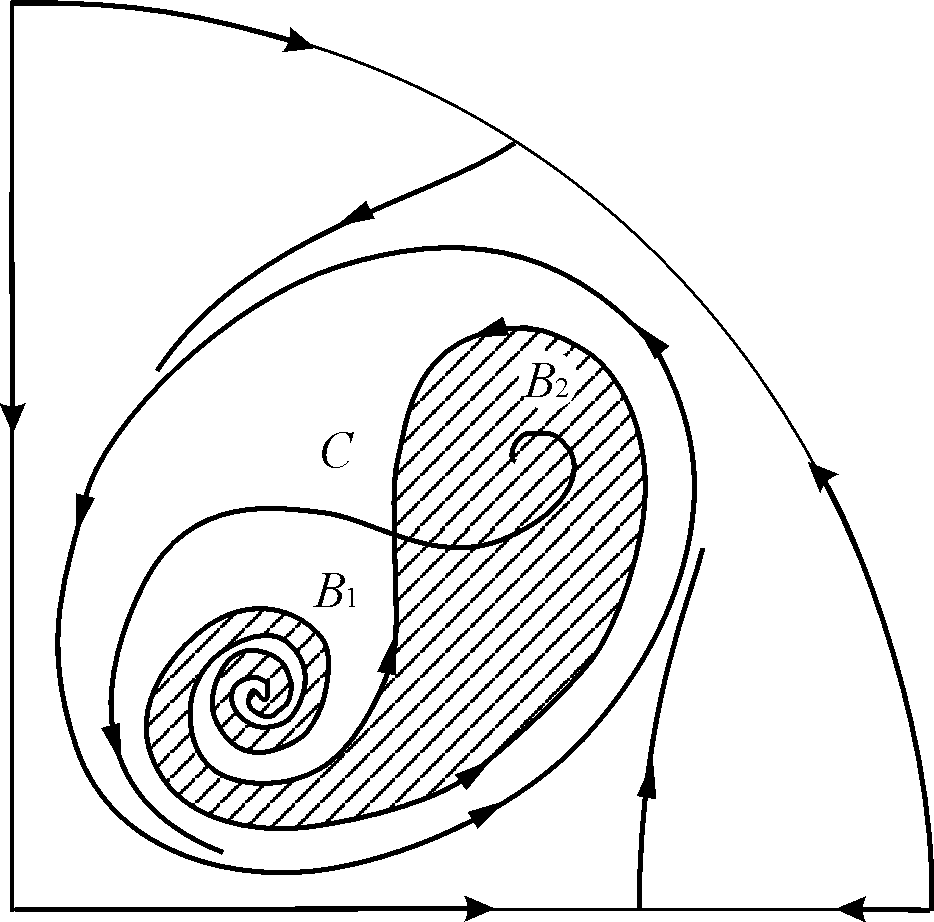
Рис.9.10. Фазовый портрет системы 9.18 для параметрической области 6. Область притяжения В2 заштрихована
На параметрическом портрете (9.7) имеются 22 различные бифуркационные границы, которые образуют 7 различных типов бифуркаций. Их изучение позволяет выявить возможные типы поведения системы при изменении ее параметров. Например, при переходе из области 1 в область 3 происходит рождение малого предельного цикла, или мягкое рождение автоколебаний вокруг единственного равновесия В. Аналогичное мягкое рождение автоколебаний, но вокруг одного из равновесий, а именно B1, происходит при пересечении границы областей 2 и 4. При переходе из области 4 в область 5 устойчивый предельный цикл вокруг точки B1 “лопается” на петле сепаратрис и единственной притягивающей точкой остается равновесие B2 и т.д.
Особый интерес для практики представляет, конечно, выработка критериев близости системы к бифуркационным границам. Действительно, биологам хорошо известно свойство “буферности”, или “гибкости”, природных экологических систем. Этими терминами обычно обозначают способность системы как бы поглощать внешние воздействия. Пока интенсивность внешнего воздействия не превышает некоторой критической величины, поведение системы не претерпевает качественных изменений. На фазовой плоскости это соответствует возвращению системы в устойчивое состояние равновесия или на устойчивый предельный цикл, параметры которого не сильно отличаются от первоначального. Когда же интенсивность воздействия превышает допустимую, система “ломается”, переходит в качественно иной режим динамического поведения, например просто вымирает. Это явление соответствует бифуркационному переходу.
Каждый тип бифуркационных переходов имеет свои отличительные особенности, позволяющие судить об опасности такого перехода для экосистемы. Приведем некоторые общие критерии, свидетельствующие о близости опасной границы. Как и в случае одного вида, если при уменьшении численности одного из видов происходит “застревание” системы вблизи неустойчивой седловой точки, что выражается в очень медленном восстановлении численности к начальному значению, значит, система находится вблизи критической границы. Индикатором опасности служит также изменение формы колебаний численностей хищника и жертвы. Если из близких к гармоническим колебания становятся релаксационными, причем амплитуда колебаний увеличивается, это может привести к потере устойчивости системы и вымиранию одного из видов.
Итак, мы рассмотрели автономные непрерывные математические модели, описывающие взаимодействие двух видов. Сделаем некоторые выводы. При моделировании биоценоза из двух видов система Вольтерра (9.1) дает возможность для описания устойчивого сосуществования видов в условиях конкуренции, симбиоза и хищничества (паразитизма). При попытке описать устойчивые колебания численности видов мы сталкиваемся с трудностями. Система уравнений (5.17), описывающая взаимодействия хищник-жертва без учета самоограничения численности популяций и имеющая особую точку типа центр, ‑ негрубая и, следовательно, неустойчива к случайным флуктуациям численности. Предельных же циклов, являющихся фазовыми траекториями устойчивых автоколебаний, система типа Вольтерра (9.1) иметь не может. Для получения предельных циклов в модельных системах приходится выходить за рамки гипотез Вольтерра и учитывать более тонкие эффекты взаимодействия между видами. Правые части уравнений при этом становятся существенно нелинейными.
Дальнейшее углубление математической теории взаимодействия видов идет по линии детализации структуры самих популяций и учета временных и пространственных факторов.