

13. Лимбическая система, структуры и функции.
Под лимбической системой понимают ряд корковых и подкорковых структур, обеспечивающих организацию мотивационно-эмоциональных реакций, процессы памяти и обучения. К структурам лимбической системы (limbus – край) относятся: гипоталамус, передневентральное ядро таламуса, поясная извилина (сингулярная кора), гиппокамп, мамиллярные тела гипоталамуса, свод, перегородка, орбито-фронтальная кора, расположенная на базальной части лобной доли мозга, и миндалина.

Афферентные импульсы поступают в лимбическую систему от различных областей коры головного мозга, от обонятельных рецепторов, гипоталамуса и ретикулярной формации. После обработки полученной информации лимбическая система запускает вегетативные и соматические поведенческие реакции, обеспечивающие приспособление организма к меняющимся условиям среды.
Через гипоталамус большинство лимбических структур объединено в целостную систему, регулирующую мотивационно-эмоциональные реакции, обеспечивающие адаптивное поведение человека, построенное на основе доминирующей биологической мотивации. Предполагают, что внешние и внутренние стимулы возбуждают мотивационные центры гипоталамуса, который в свою очередь активирует гиппокамп и фронтальные отделы коры головного мозга, связанные с прогнозированием вероятности достижения положительного результата. Установлено месторасположение в различных отделах гипоталамуса центров положительных и отрицательных эмоций.
Не менее важную роль в формировании эмоций играют миндалина и поясная извилина. Поясная извилина отвечает за интеграцию различных систем мозга, участвующих в формировании эмоций. Миндалина участвует в нескольких видах эмоционального поведения: агрессии, страхе, отвращении, материнском поведении, активируя соответствующие участки гипоталамуса и других структур ствола мозга. Благодаря сопоставлению потребностей с имеющимися стимулами и с энграммами извлеченными из памяти, в миндалине формируется окраска этих стимулов (вкусно-невкусно, знакомонезнакомо), что приводит к определению мотивации, подлежащей удовлетворению. Выбор доминирующей потребности миндалина осуществляет путем взвешивания конкурирующих эмоций. Раздражение центрального ядра миндалины ведет к изъязвлению желудка, т.к. через соответствующие проекции она вовлечена в реализацию гормонов стресса.
Лимбическая система участвует в обеспечении процессов памяти и обучения. Существует мнение, что перевод информации из кратковременной в долговременную память происходит во сне. Важную роль при этом играет гиппокамп. Во время сна гиппокамп манипулирует со следами памяти, обнаруживая общие элементы в различных комплексах стимулов, и не допускает консолидации в памяти случайных следов.

14. Строение и функции отделов автономной (вегетативной) нервной системы: симпатического, парасимпатического, метасимпатического. Энтеральная система. Роль отделов центральной нервной системы в регуляции вегетативных функций.
Часть центральной нервной системы (ЦНС), регулирующая работу внутренних органов и постоянство внутренней среды организма (гомеостаз), называется вегетативной или автономной нервной системой. Деятельность вегетативной нервной системы только в малой степени зависит от коры больших полушарий, т.е. работа совершается автоматически, минуя сознание.
Характерной особенностью эфферентных путей, входящих в рефлекторные дуги вегетативных рефлексов, является их двухнейронное строение. Один нейрон находится в ЦНС, другой – в ганглиях или иннервируемом органе. Волокна автономной системы отличаются сравнительно низкой возбудимостью и скоростью проведения импульсов. Автономная нервная система подразделяется на симпатический, парасимпатический и метасимпатический отделы.

Центральные нейроны симпатического отдела локализованы в боковых рогах спинного мозга от 1-го грудного до 2-го поясничного сегмента. Вторые нейроны симпатического отдела располагаются в превертебральных вегетативных ганглиях. Характерной особенностью симпатических волокон является короткий преганглионарный и длинный постганглионарный пути (рис. 32). В преганглионарных волокнах в основном секретируется медиатор ацетилхолин, в посганглионарных – норадреналин. Симпатические волокна иннервируют все органы и ткани организма, включая ЦНС и сенсорные рецепторы. Симпатическая часть автономной нервной системы способствует интенсификации деятельности организма, особенно в экстремальных условиях. К симпатическим рефлексам, обеспечивающим активную деятельность, относятся рефлексы расширения бронхов, усиления и учащения сердечных сокращений, расширения сосудов сердца и легких при одновременном сужении сосудов кожи и органов брюшной полости, выброс депонированной в печени и селезенке крови, расщепление гликогена печени, усиление деятельности желез внутренней секреции и потовых желез.
Центральные парасимпатические нейроны расположены в структурах ствола мозга (среднем и продолговатом мозге) и крестцовом отделе спинного мозга. Вторые нейроны находятся в парасимпатических ганглиях, расположенных недалеко от иннервируемых органов либо внутри них. От нервных клеток парасимпатических ганглиев идут короткие постганглионарные волокна, тогда как преганглионарные волокна обычно длинные. В нервных окончаниях пре- и постганглионарных нейронов секретируется медиатор ацетилхолин. Пусковые влияния парасимпатической нервной системы связаны с рефлексами сужения зрачка, бронхов, сосудов сердца, включение деятельности пищеварительных желез и т.д. В целом деятельность парасимпатического отдела ВНС направлена на текущую регуляцию функционального состояния и восстановление нарушенного во время активности организма гомеостаза (постоянства внутренней среды). Таким образом, симпатический и парасимпатический отделы оказывают на ткани, органы и функции антагонистическое, но содружественное влияние, что обеспечивает более точную регуляцию. Поэтому к большинству внутренних органов подходят два нерва: симпатический и парасимпатический. Парасимпатической иннервации не имеют только скелетные мышцы, матка, мозг, подавляющее большинство кровеносных сосудов (кожи, органов брюшной полости, мышц), органы чувств и мозговое вещество надпочечников.
Метасимпатические нейроны располагаются в стенках внутренних органов, обладающих моторной активностью: в желудке, кишечнике, мочевом пузыре, сердце, бронхах, шейке матки. Плотность их расположения во внутренних органах очень высока, например, на 1 см2 кишечника приходится около 20 тыс. нейронов. Метасимпатическая нервная система содержит все элементы рефлекторной дуги: афферентный, вставочный и эфферентный нейроны, поэтому может функционировать и после перерезки симпатических и парасимпатических нервов. Все отделы вегетативной нервной системы подчинены вегетативным центрам головного мозга, расположенным в продолговатом мозге, заднем мозге (мост и мозжечок), среднем мозге, промежуточном мозге (гипоталамус), базальных ганглиях, лимбической системе, ретикулярной формации и коре больших полушарий. Кора головного мозга, в частности премоторная зона, получает импульсы от всех внутренних органов и с помощью ВНС оказывает на них влияние. Особое значение для регуляции вегетативных функций имеет гипоталамус



Энтера́льная не́рвная систе́ма — часть вегетативной нервной системы, регулирующая работу гладких мышц внутренних органов, обладающих сократительной активностью.
Нервные сплетения, составляющие энтеральную нервную систему, располагаются в оболочках полых органов желудочно-кишечного тракта (пищевод, желудок, тонкая и толстая кишка, выводящие желчные и панкреатические протоки, сфинктер Одди и др.), мочевыделительной системы (лоханки и чашечки почек, мочеточники, мочевой пузырь и др.). Важную роль в ритмической моторной активности этих органов играют такие элементы энтеральной нервной системы, как двигательные нейроны и клетки — водители ритма.

15. Виды мышечной ткани, строение мышечной клетки. Механизм сокращения и расслабления скелетной мышечной ткани. Теория скольжения нитей. Роль саркоплазматического ретикулума и ионов кальция в сокращении. Особенности сокращения гладкой мышечной ткани.
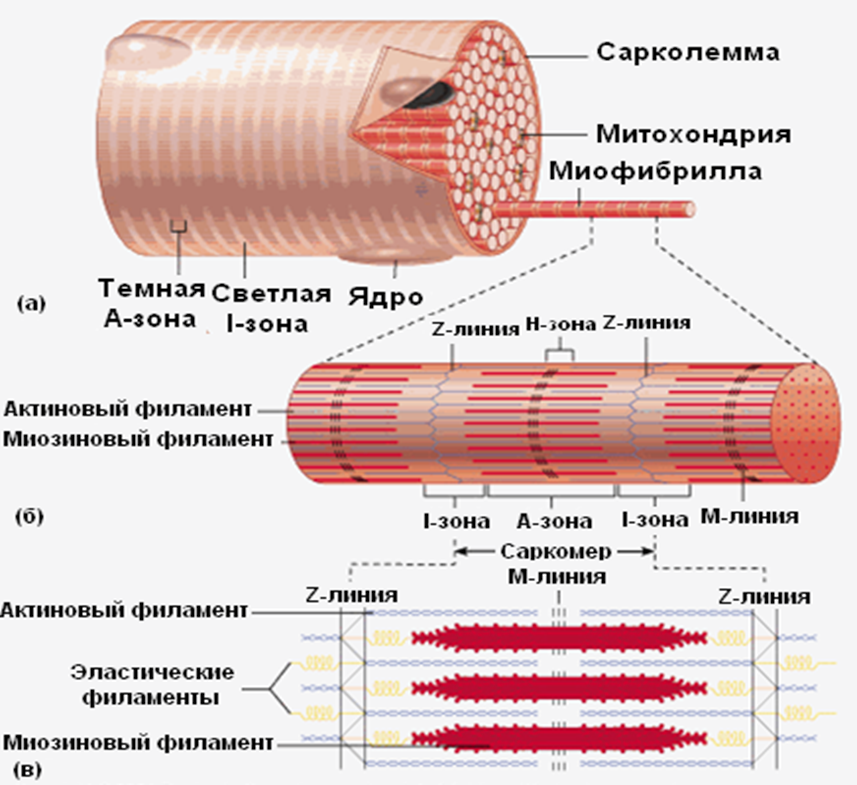
Ультраструктура сократительного аппарата. Мышечные волокна, из которых построены скелетные мышцы, представляют собой гигантские многоядерные клетки, достигающие длины 50 мм и толщины 50 мкм. Основную часть их цитоплазмы занимают миофибриллы – цилиндрические белковые элементы с характерными регулярно чередующимися поперечными светлыми и темными полосками. Поперечнополосатая исчерченность миофибрилл обусловлена наличием сократительных белков – тонких нитей актина и толстых нитей миозина, скомпонованных в отдельные отсеки длиной 2,5 мкм – саркомеры.
Саркомеры ограничены друг от друга перегородками, называемыми Z-дисками, иначе говоря, Z-мембранами, к которым крепятся актиновые нити, образующие светлый (изотропный) I-диск. В центре саркомера расположены толстые миозиновые нити, образующие темный (анизотропный, из-за двойного лучепреломления в поляризованном свете) А-диск длиной 1,6 мкм. Чуть более светлая полоса посередине А-диска – это место прикрепления нитей миозина к М-мембране (узкой темной полоске). На уровне Zмембран к каждому саркомеру спускается поперечная Т-трубочка, являющаяся продолжением продольной плазматической мембраны. Эти трубочки контактируют с сетью продольных трубочек и цистерн саркоплазматического ретикулума, который содержит ионы Са2+.
Молекула миозина состоит из шести полипептидных цепей: двух тяжелых и двух пар легких. Толстые миозиновые филаменты образуются путем переплетения многочисленных хвостов миозина в пучок, на поверхность которого выступают головки миозина, обеспечивающие контакт и перемещение толстых филаментов относительно тонких актиновых филаментов. Головки миозина соединены с телом миозиновой нити с помощью шейки, которая может изгибаться. На кончике головки имеется карман, где находится одна молекула АТФ. АТФ-азная активность головки проявляется только в присутствии актина.
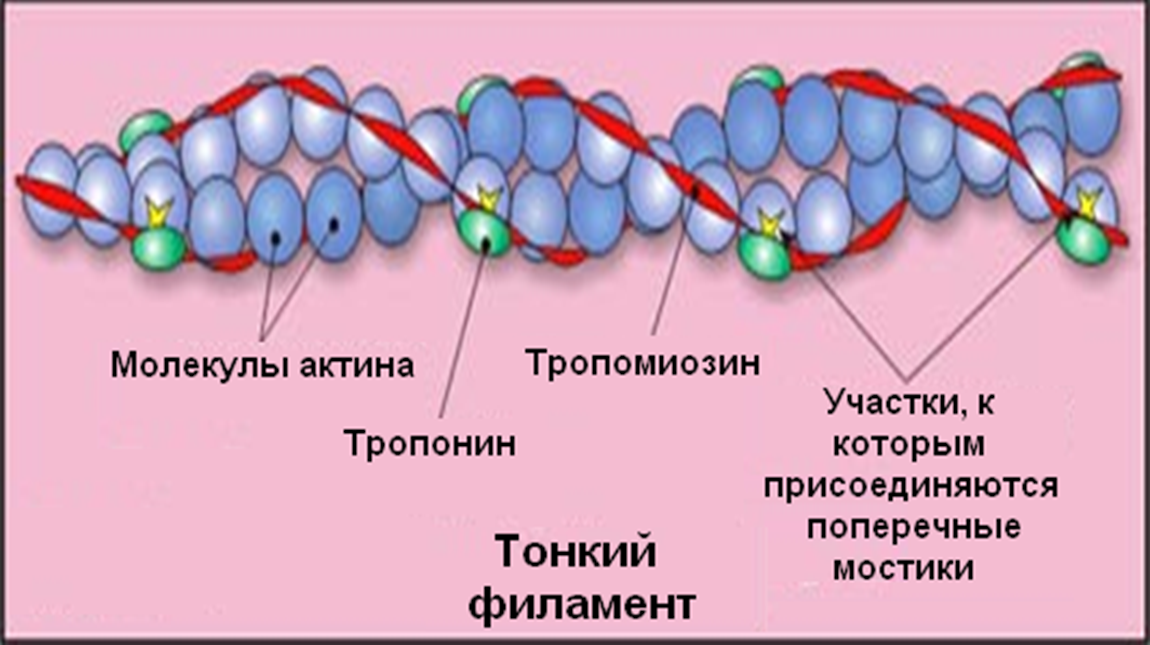
Актиновый филамент представляет собой спираль из мономеров актина, которые состоят из одной полипептидной цепи, образованной 375юаминокислотными остатками. Жесткость актиновым филаментам придают молекулы тропомиозина длиной 41 нм, состоящие из двух скрученных полипептидных цепей (по 284 аминокислотных остатка), связанных с молекулами актина. С тропомиозином контактирует молекула тропонина. В состоянии покоя тропонин связан с актином, а тропомиозин занимает место, где должен быть контакт с головками миозина, поэтому взаимодействия актина с миозином не происходит.
Механизм сокращения и расслабления мышечного волокна
В момент генерации ПД, ионная природа которого в фазных мышечных волокнах аналогична таковому нервного волокна, происходит распространение ПД вдоль продольной плазматической мембраны, включая поперечные трубочки, контактирующие с цистернами саркоплазматического ретикулума. В результате открываются кальциевые каналы, по которым кальций выходит в межфибриллярное пространство. Здесь он связывается с молекулами тропонина, после чего тропомиозин освобождает участок связывания актина с миозином, что дает возможность для осуществления цикла мышечного сокращения.
Сокращение происходит согласно распространенной модели «скользящих нитей» А. Хаксли (1971). Головки миозина образуют поперечные мостики, цепляющиеся к актиновой нити под углом 90°. Тут же происходит активация АТФ-азной активности и как следствие гидролиз АТФ до АДФ и фосфата с выделением порции энергии. Далее головка миозина освобождает фосфат и связывается с актином. Затем головка миозина претерпевает сильные конформационные изменения, в результате чего совершает крутящий момент (гребок) и проталкивает актиновую нить примерно на 10 нм. Если рядом с образовавшимся мостиком имеется молекула АТФ, то она встраивается в вершину мостика и обеспечивает отрыв головки миозина от актиновой нити. Затем опять происходит гидролиз АТФ, и цикл повторяется. Миозиновая головка вновь прикрепляется к актиновой нити, но уже в другом месте, до тех пор, пока в среде будет достаточно кальция.
Расслабление мышцы происходит в результате снижения уровня кальция, т.к. мембраны продольных трубочек саркоплазматического ретикулума начинают откачивать кальций назад. Таким образом, миозиновые головки как бы «шагают» по актиновому филаменту, продвигая его относительно себя.
