

Энергетические процессы в митохондриях. Строение и молекулярная организация митохондрий
Митохондрии, число которых в клетке составляет несколько сотен (например, в гепатоцитах 1700), занимают значительную часть цитоплазмы почти всех эукариотических клетках (в гепатоцитах около 22%). Обычно митохондрии скапливаются вблизи мест интенсивного потребления АТФ - например, в сердечной мышце они располагаются между миофибриллами. Митохондрии достаточно велики (до 1 мкм и больше), и их можно увидеть в световой микроскоп, в виде жестких вытянутых, похожих на бактерии цилиндров. Однако микрокиносъемка живых клеток позволяет увидеть, что митохондрии - необыкновенно подвижные и пластичные образования, которые постоянно изменяют свою форму и даже сливаются друг с другом, а затем вновь разделяются. Детальное строение митохондрий, их ультраструктура была выяснена только с помощью электронной микроскопии. Оказалось, что каждая митохондрия окружена двумя бислойными мембранами, играющими ключевую роль в ее функции. Мембраны образуют два изолированных митохондриальных компартмента: внутренний матрикс и значительно более узкое межмембранное пространство. Основная рабочая часть митохондрии - это матрикс и окружающая его внутренняя мембрана. Площадь внутренней мембраны в 4,5 раза больше чем внешней, т.к. она образует многочисленные складки - кристы, увеличивающие общую поверхность. На внутренней мембране локализована система окислительного фосфорилирования, включающая АТФ-синтетазу и дыхательную цепь (син. электрон-транспортная цепь митохондрий). Важнейшим физиологическим свойством внутренней мембраны является непроницаемость ее для ионов.
Матрикс представляет собой высококонцентрированнай раствор сотен различных ферментов, в том числе ферментов цикла трикарбоновых кислот. В матриксе находится митохондриальная ДНК и митохондриальные рибосомы.
Цикл трикарбоновых кислот
В большинстве клеток и тканей аэробных организмов, образующаяся в результате гликолиза пировиноградная кислота поступает в матрикс митохондрий, где она декарбоксилируется, т.е. от нее отщепляется молекула углекислого газа, а оставшаяся двухуглеродная ацетильная группа соединяется с коферментом А (СоА), образуя ацетил-СоА. Энергия, выделяющаяся в результате этой реакции, расходуется на перевод специальной молекулы - переносчика водорода из окисленной в восстановленную форму. Переносчик водорода называется никотинамидадениндинуклеотид и обозначается сокращенно NAD (НАД) или NAD+ (окисленная форма) и NADH (НАДН) или NADH + H+ (восстановленная форма). Таким образом, молекулы NAD и сходного с ним по структуре и функции флавинадениндинуклеотида, сокращенно FAD, подобно фосфагенам способны запасать и переносить энергию, но не в виде макроэргической фосфатной связи, а в виде активированных атомов водорода. Образовавшийся из пировиноградной кислоты ацетил-СоА подвергается дальнейшему окислению в цикле трикарбоновых кислот. В результате первой реакции цикла (Рис. ) из ацетил-С и щавелевоуксусной кислоты (в водной среде существует в виде иона - оксалоацетата) содержащей 4 атома углерода при участии фермента цитратсинтазы образуется трикарбоновая, шестиуглеродная лимонная кислота (цитрат). Поэтому цикл трикарбоновых кислот называют иногда цикл лимонной кислоты. Затем лимонная кислота превращается в изолимонную (изоцитрат). В результате следующей, реакции декарбоксилирования шестиуглеродная молекула изолимонной кислоты превращается в пятиуглеродную α-оксоглутаровую кислоту (α-оксоглутарат) и происходит восстановление NAD в NADH. Далее α-оксоглутаровая кислота декарбоксилируется до янтарной (сукцинат). При этом выделяющаяся энергия не рассеивается в виде тепла, а затрачивается на восстановление еще одной молекулы NAD в NADH и образование промежуточного продукта с высокоэнергетической фосфатной связью, который обеспечивает фосфорилирование гуанозиндифосфата (GDP) в гуанозинтрифосфат (GTP).
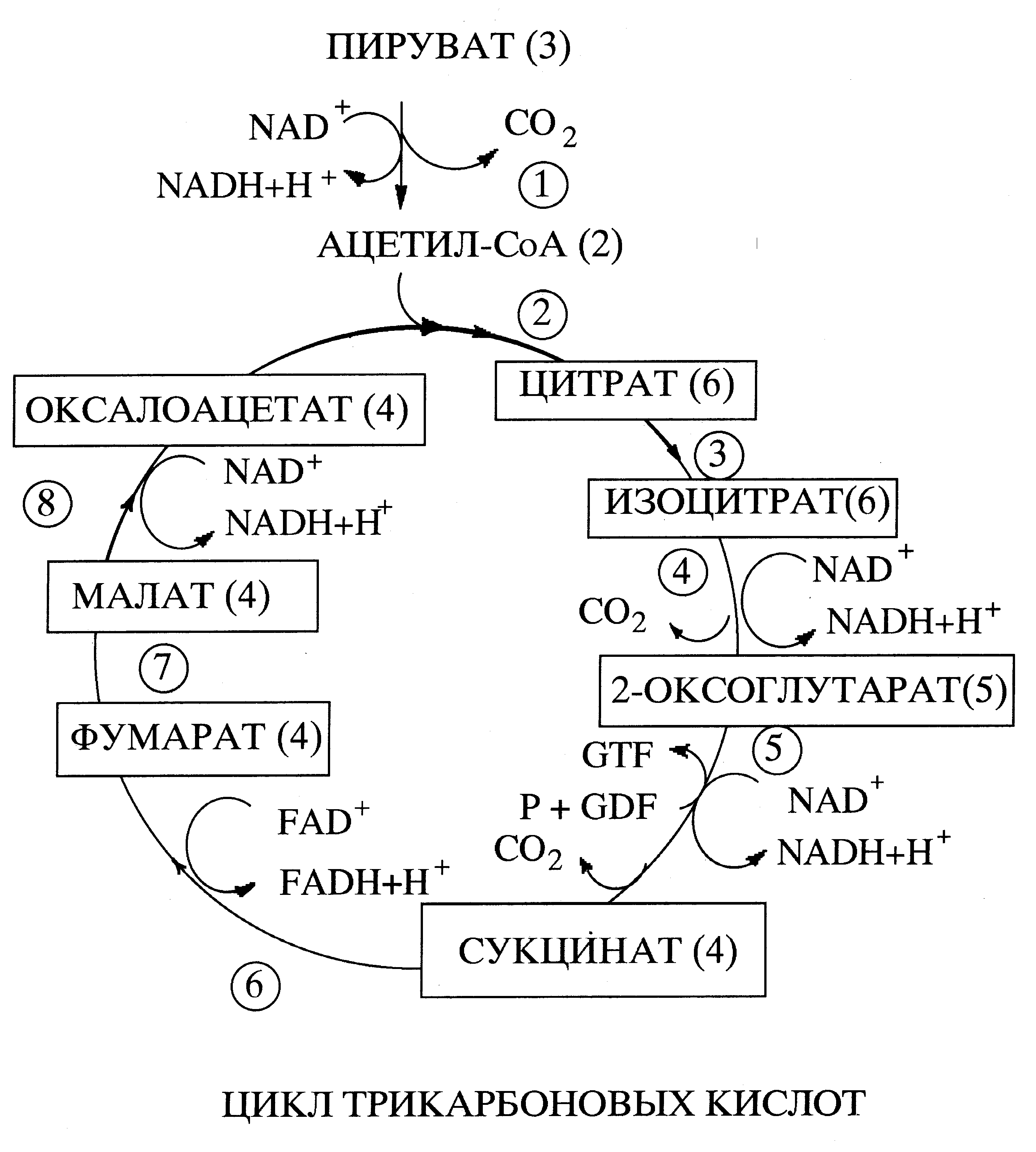
В результате трех следующих реакций янтарная кислота последовательно превращается в фумаровую кислоту (фумарат), яблочную кислоту (малат) и щавелевоуксусную и восстановливается по молекуле FAD и NAD.
Таким образом, ферментативные реакции цикла трикарбоновых кислот протекают так, что субстрат первой реакции - щавелевоуксусная кислота является и продуктом последней восьмой реакции, т.е. регенерируется после каждого оборота цикла и процесс запускается вновь. В результате элементарного цикла два углеродных атома, имевшихся в ацетил-СоА, превращаются в СО2. Циклический характер последовательности реакции окисления ацетил-СоА и ее основные этапы установил в 1937 году английский биохимик немецкого происхождения, лауреат Нобелевский премии (1953) Ханс Кребс. Поэтому процесс окисления ацетил-СоА имеет еще одно название - цикл Кребса. Благодаря тому, что окисление совершается постепенно, через серию относительно небольших изменений свободной энергии, выделяющаяся энергия не рассеивается в виде бесполезного тепла, а затрачивается на восстановление трех молекул NAD в NADH и одной молекулы FAD в FADH и образование высокоенергетической фосфатной связи. Эта связь образуется в результате фосфорилирования гуанозиндифосфата в гуанозинтрифосфат. Последний легко обменивается фосфатной группой с АДФ с образованием АТФ.
Следует иметь в виду, что источником ацетил-CoA для окисления в ЦТК, наряду с гликолизом являются и другие биохимические процессы протекающие в клетках, в частности окисление аминокислот и жирных кислот. Четыре молекулы NADH, включая и образовавшуюся при окислении пирувата в ацетил-СоА, и молекула FADH, поступают в дыхательную цепь, где переносимые ими атомы водорода окисляются молекулярным кислородом до воды. Выделяемая при этом энергия затрачивается на синтез АТФ из АДФ и неорганического фосфата в результате сложнейшего, во многом неясного процесса, получившего название окислительного фосфорилирования.