

Теоретична частина:
Матричні біосинтези включають:
-
Реплікацію (подвоєння ДНК),
-
Репарацію (процеси виправлення ушкоджень, які виникають у ДНК),
-
Транскрипцію (біосинтез іРНК, тРНК, рРНК),
-
Трансляцію (біосинтез білка).
Основна схема передачі генетичної інформації:
транскрипція трансляція


 ДНК
іРНК
білок
ДНК
іРНК
білок
зворотна
Реплікація (у ядрі клітини) – синтез дочірньої молекули ДНК на матриці материнської молекули ДНК, заснований на принципі компліментарності азотистих основ (А=Т, Г ≡Ц), відбувається у S-фазу клітинного циклу.
Механізм реплікації – полуконсервативний. У результаті реплікації утворюється 2 нові молекули ДНК, у кожній із них – 1 ціпок „материнський”, 2 – „дочірній”.
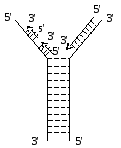
У
ДНК була виявлена активна У-подібної
форми область, яка має назву реплікаційна
вилка. У цих двох антипаралельних ціпках
синтез дочірніх ціпків йде у напрямку
5’-3’. При цьому один ціпок будується
неперервно (ведучій ціпок), а другий
фрагментами (запізнюючий ціпок) по
1000-2000 нуклеотидів (фрагменти Оказаки)
ферментом ДНК-полімеразою. Ці
фрагменти в
подальшому сполучаються
в довгий
дочірній
ціпок ДНК-лігазою.
Кожний фрагмент та ведучий ланцюг
починаються із синтезу затравки
(праймер), яка
будується з рибонуклеотидів. Синтез іде за допомогою ДНК-праймази. Затравка складається із 10 рибонуклеотидів та її ще називають РНК-затравкою.
Затравки
синтезуються з визначеним інтервалом
на матриці для відстаючого ланцюга.
Така Рнк-затравка
далі нарощується Днк-полімеразою,
але вже дезоксирибонуклеотидами доти,
доки вона не досягне Рнк-затравки
попереднього фрагменту. Потім РНК-затравки
вирізаються
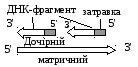
і замість них ДНК-полімераза вбудовує дезоксирибонуклеотиди.
На ведучому та запізнюючому ланцюгах діють 2 однакові ДНК-полімерази.
Транскрипція – біосинтез молекул РНК на матриці ДНК, який локалізований в ядрі клітини, йде постійно і незалежно від циклу клітини. Ферментом транскрипції є РНК-полімераза. В евкаріот різні види РНК синтезуються різними РНК-полімеразами. У прокаріот – однією. Матрицею для синтезу ірнк служить певна ділянка одного з ланцюгів ДНК, який має назву ген, або цистрон. Молекула ірнк будується за принципом комплементарності й несе в собі генетичну інформацію про структуру одного певного білка.
РНК-полімераза приєднується до специфічної послідовності - Днк-промотору, який містить старт-сигнал для синтезу РНК. Приєднавшись до промотору, Рнк-полімераза розкручує визначену ділянку подвійної спіралі, оголюючи таким способом нуклеотиди на короткому відрізку кожного з двох ланцюгів ДНК. Один з них служить матрицею для комплементарного спарювання основ ДНК із основами мономерів – рибонуклеозидтрифосфатів. Полімераза з'єднує між собою два перші мономери і таким чином починає синтез РНК. Просуваючи уздовж ДНК, Рнк-полімераза розкручує спіраль, оголюючи нову ділянку матриці. Ланцюг РНК нарощується в напрямку 5'→3'.
Процес подовження ланцюга відбувається доки фермент не зустріне на своєму шляху специфічну нуклеотидну послідовність у ланцюзі ДНК – сигнал термінації транскрипції (стоп-сигнали). Полімераза відокремлюється від ДНК і від РНК. Під час просування ферменту уздовж матричного ланцюга в його активному центрі утворюється подвійна спіраль ДНК-РНК. Вона дуже коротка, тому що за молекулою Рнк-полімерази негайно відновлюється спіраль ДНК-РНК, а РНК – витісняється.
Завжди транскрибується (копіюється) тільки один із двох ланцюгів.
Дозрівання РНК (процесинг) відбувається в ядрі поза матриці під впливом РНК-аз, потім зріла РНК виходить з ядра в цитоплазму. іРНК переміщується на рибосому, де вона служить матрицею для синтезу білка. Таким чином, генетична інформація, закодована в ДНК, переводиться (транслюється) рибосомою в послідовність амінокислот, які утворюють різноманітні функціональні і структурні білки клітини.
Більші частини амінокислот, які надійшли в тканини разом з током крові, використовуються для біосинтезу тканьових білків. Вивчення механізму біосинтезу білка - це та область сучасної біохімії, яка найбільше інтенсивно розробляється і в якій досягнуті останнім часом значні успіхи.
У біосинтезі білка беруть участь ДНК і всі три типи РНК - інформаційна, транспортна і рибосомна. Етапами трансляції є: ініціація, елонгація і термінація.
Синтез кожного з тисяч різноманітних білків клітини кодується специфічною ірнк. Звідси стає зрозумілим велике (до декількох тисяч) число видів ірнк, які істотно відрізняються за молекулярною масою (від 25 тис. до 1 млн. ) і нуклеотидним складом.
Попередній етап – утворення активованих амінокислот за участю ферменту аміноацил-т-РНК-синтетази, 1 молекули АТФ, Mg2+, 61 вида тРНК. Фермент високо специфічний як у відношенні однієї певної амінокислоти, так і у відношенні трнк.
Mg2+
А мінокислота
+ тРНК + АТФ Аміноацил-тРНК + АМФ +
пірофосфат.
мінокислота
+ тРНК + АТФ Аміноацил-тРНК + АМФ +
пірофосфат.
Транспортна РНК приєднує активовану амінокислоту до так званого акцепторного кінця і переносить її до рибосоми – білоксинтезуючій частці клітини, яка складається із двох нерівних за розміром субчастиць (30 і 50 S у прокаріот; 45 і 60S у евкаріот), здатних до дисоціації і реасоціації.
Особливістю будови трнк є наявність у її середній петлі антикодону - трьох нуклеотидів (триплету) - певного складу для кожної амінокислоти. Антикодони трнк комплементарні відповідним кодонам (триплетам) інформаційної РНК і являють собою точні копії триплетів тих ділянок ДНК (генів), які були використані при транскрибуванні інформаційної РНК. У процесі синтезу білка відбувається взаємодія антикодонів транспортної РНК із комплементарними кодонами інформаційної РНК.
Ініціацією називають початок синтезу білка. На рибосомі відбувається приєднання спеціальної ініціаторної транспортної РНК, завжди зв'язаної у евкаріот із залишком амінокислоти метіоніну (метіоніл-трнк), а в прокаріот - з N-формілметіоніном, до ініціюючого кодона інформаційної РНК, яким є триплет АУГ (аденін, урацил, гуанін). Утворення ініціюючого комплексу інформаційної РНК із метіоніл-трнк відбувається на меншій за розміром субчастці, яка потім зв'язується з більшою субчасткою.
Елонгація – подовження поліпептидного ланцюга шляхом почергового приєднання амінокислотних залишків до С-кінця зростаючого поліпептидного ланцюга. Після утворення чергового пептидного зв'язку відбувається транслокація, тобто переміщення на рибосомі ірнк і пептидил-трнк.
Термінація – закінчення синтезу поліпептидного ланцюга і його відділення від рибосоми, яка знову дисоціює на субчастки. Сигналом термінації служать три специфічних кодони ірнк (УАА, УГА, УАГ).
У клітині з молекулою ірнк зв'язано кілька рибосом. Цю систему називають полірибосомою, або полісомою. Таким чином, на одній молекулі ірнк іде одночасно синтез декількох поліпептидних ланцюгів, що підвищує ефективність використання ірнк і значно прискорює синтез білка. На утворення одного пептидного зв'язку витрачається не менш трьох макроергічних фосфатних зв'язків. Регуляція біосинтезу білка в клітині здійснюється як на рівні транскрипції, так і на рівні різних етапів трансляції. Стероїдні гормони діють на рівні геному, стимулюючи синтез специфічних інформаційних РНК.
Окремі амінокислоти, крім їх участі в побудові молекули білка, використовуються клітинами організму для синтезу ряду біологічно активних азотистих небілкових речовин. Так, наприклад, з амінокислоти тирозину утворюється гормон щитовидної залози - тироксин і гормони мозкового шару наднирників – адреналін і норадреналін. Гліцин, аспарагінова кислота і глютамін беруть участь у синтезі пуринових і піримідинових основ. У біосинтезі гема (простетичної групи гемоглобіну) у якості одного з основних субстратів використовується гліцин. Із гліцину, аргініну і метіоніну синтезується креатин, який, приєднуючи фосфорильну групу АТФ, перетворюється в багату енергією сполуку – фосфокреатин, яка грає роль енергетичного "депо" у м'язах. Стандартна вільна енергія гідролізу зв'язку -N~Р- у фосфокреатині при фізіологічних значеннях рн становить приблизно 9ккал/моль. Окремі амінокислоти беруть участь у біосинтезі пептидних гормонів - вазопресину, окситоцину, брадікініну - і в синтезі цілого ряду біологічно активних коротких пептидів, таких як трипептид глютатіон (- глютамінілцистеїнілгліцин) і дипептиди карнозин (-аланілгістидин) і анзерин (-аланілметилгістидин). У рослинах ароматичні амінокислоти служать попередниками багатьох алкалоїдів. З тирозину утворюються алкалоїди - морфін, кодеїн, папаверин, а із триптофану - індолілоцтова кислота - гормон росту рослин.