

1.6.Мітохондрії
Мітохондрія - двомембранна органела евкаріотичної клітини, що забезпечує її енергією за рахунок процесів аеробного окислення органічного субстрату.
Внутрішня мембрана мітохондрій, яка складається переважно з білків (70%), утворює виступи - кристи. Середовище мітохондрії - матрикс, містить ферменти циклу трикарбонових кислот, кільцеву ДНК, РНК, рибосоми. Завдяки цьому мітохондрії, як і пластиди, є напівавтономними органелами. Вони розмножуються поділом і мають власний білоксинтетичний апарат. На внутрішній мембрані відбуваються процеси окисного фосфорилювання, внаслідок чого виникає електрохімічний потенціал, що витрачається на синтез АТФ.
1.7.Вакуолярна система
До одномембранних органел, які різняться перш за все функціонально, але подібні в структурному відношенні, належать апарат Гольджі (АГ), ендоплазматичний ретикулум (ЕПР), вакуолі, лізосоми і т.п. Інколи їх об'єднують під загальною назвою вакуолярна система. Фактично вона являє систему мембран, які поділяють протопласт на відмінні функціональні частини, де иідбувається протікання різних метаболічних процесів: синтез (ЕПР), перенесення (АГ), перетворення (ЕПР, АГ), руйнування (лічосоми) речовин, переведення їх в нерозчинних стан (вакуолі) і т.п. Лише завдяки системі цих мембран в морфологічно єдиній цитоплазмі можливий одночасний перебіг різноманітних життєвих процесів. Вакуолярна система динамічна, між окремими її компонентами відбувається кругообіг мембран, внаслідок чого постійно змінюється ємність перерахованих органел залежно від потреб, а частина мембран однієї органели може стати складовою іншої та навпаки.
Апарат Гольджі (АГ) - багатоярусна система плоских порожнистих дископодібних цистерн, яка бере участь у транспорті та хімічній модифікації речовин.
До складу АГ входять диктіосоми та пухирці. Розрізняють секреторний та регенеративний бік апарату Гольджі. На регенеративному боці поверхня диктіосом збільшується за рахунок злиття з пухирцями, що відділяються переважно від гладенького ЕПР, на секреторному боці від поверхні диктіосом відшнуровуються пухирці. Мембранна система апарату Гольджі перебуває в постійому перетворенні, разом з цим перетворюються і речовини, що знаходяться у внутрішній порожнині диктіосом. Таким чином певні речовини транспортуються до апарату Гольджі, здебільшого з ними відбуваються зміни за рахунок впливу ферментів, що також можуть надходити до АГ з іншими пухирцями, в процесі дозрівання речовини попадають з регенеративного на секреторний бік, звідки і відшнуровуються з новоутвореними пухирцями та рухаються в напрямку місць використання, а зменшення мембранної поверхні компенсується за рахунок нових пухирців, що вбудовуються в диктіосоми на регенеративному кінці. Таким чином апарат Гольджі бере важливу участь в процесах хімічного перетворення та транспорту речовин, зокрема у рослин він секретує назовні компоненти клітинної стінки.
Ендоплазматичний ретикулум (ЕПР) — мембранна система, що забезпечує компартментацію евкаріотичних клітини, бере важливу участь у синтетичних процесах і утворенні клітинних мембран.
Ендоплазматичний ретикулум буває агранулярним (гладеньким) та гранулярним (шорстким). На мембрані гранулярного ЕПР розташовуються субодиниці рибосом, утворюючи комплекси - полірибосоми. Гранулярний ЕР забезпечує синтез і транспорт білків. На рибосомах формується первинна структура білка, тобто власне поліпептидний ланцюг, потім через канал в мембрані ЕПР він попадає в порожнину ЕПР, де від нього відщеплюється сигнальна (стартуюча) послідовність амінокислот і після цього він набуває третинної структури. Агранулярний ЕПР не має на поверхні рибосом, бере участь у синтезі ліпідів з жирних кислот на гліцерола, а також у синтезі стероїдних ліпідів. Таким чином в ЕПР синтезуються і нагромаджуються речовини, відбуваються їх хімічні перетворення і далі вони надходять переважно до апарату Гольджі, звідки і транспортуються до остаточних місць використання.
Рибосома - дрібна сферична немембранна органела з двох субодиниць (великої та малої), що складаються в основному з РНК (до 60%) та білку.
Рибосоми трапляються в ядрі, пластидах, мітохондріях, цитоплазмі, але переважна кількість їх прикріплена до мембрани ендоплазматичного ретикулума, де формуються комплекси -полірибосоми. Рибосомальна (рРНК) синтезується в ядерці, далі попередники рибосом формуються в ядрі, а остаточне збирання
субодиниць відбувається вже в цитоплазмі. Рибосоми є не лише в евкаріотичних, а і у прокаріотичних клітинах. Рибосоми прокаріот та евкаріот різняться за розмірами. Оскільки розміри їх дуже дрібні (близько 20 нм), для їх характеристики використовують умовні одиниці сведберга, що визначаються швидкістю седиментації в центрифузі. Рибосоми прокаріот, а також мітохондріальні та пластидні рибосоми евкаріот, мають розміри 708, ті що розташовуються в цитоплазмі евкаріот - 808. Основна функція рибосом - формування первинної структури білків.
Вакуоля - одномембранна сферична органела, виповнена вакуолярним соком, тобто водним розчином складного хімічного складу, відмінного від складу гіалоплазми (рідкої частини цитоплазми). Основна функція вакуолі - осмотичний гомеостаз протопласту та нагромадження певних речовин.
Від цитоплазми вакуоля відмежована тонопластом. Кількість вакуолей в клітинах буває різною. У меристематичних молодих клітинах є численні дрібні вакуолі, в зрілих клітинах вакуоля займає майже 90% їх об'єму, розташовуючись в центральній частині клітини. Крупні вакуолі є особливістю рослинних клітин, у одноклітинних тваринних організмів трапляються інші, дрібніші за розмірами скоротливі і травні вакуолі.
Оскільки концентрація вакуолярного соку відрізняється від такої в міжклітинному просторі існує градієнт, за яким речовини повинні проникати через тонопласт і рухатися в напрямку розчину меншої концентрації. Але для багатьох речовин тонопласт непроникний, тому розчинник (вода) буде переходити в розчин иищої концентрації. Так виникають осмотичні явища. Осмос - перехід молекул розчинника з області з більш високою концентрацією розчинених речовин в область з більш низькою концентрацією через напівпроникну мембрану. В процесі руху розчинника через напівпроникну мембрану виникає осмотичний тиск, який дорівнює зовнішньому тиску, який слід прикласти до мембрани, щоб припинити осмотичне надходження розчинника. Розчин, що має вищу концентрацію розчинених речовин називають гіпертонічним, однакову - ізотонічним, нижчу - гіпотонічним.
Якщо клітину занурити в гіпотонічний розчин, вода наддходить всередину клітини, збільшуючи об'єм протопласту. Виникаєсила, що діє на розрив плазмалеми. їй протистоїть механічна жорсткість клітинної оболонки, яка обумовлює тургорний тиск . Тургорний тиск - це тиск клітинної оболонки, який протидіє піску внутрішньоклітинної рідини і забезпечує збереження об'єму клітини.
Якщо клітину занурити в гіпертонічний розчин, вода викодить з клітини, об'єм протопласту зменшується, він відстає від клітинної стінки. Виникає явище плазмолізу (рис. 3.10). За невеликої в'язкості цитоплазми протопласт повністю відстає від клітинної стінки і спостерігається випуклий плазмоліз, за великої в'язкості - протопласт зберігає контакт з клітинною стінкою і спостерігається ввігнутий плазмоліз. При вміщенні в розчин нижчої концентрації відбувається деплазмоліз - повернення клітини в стан тургору в результаті додаткового надходження води
Рис.
3.10. Плазмолітичний та тургорний стан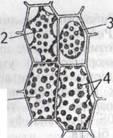
промопласту.1-тургор,2- ввігнутий плазмоліз,3- випуклий плазмоліз,4- хлоропласти.
1
За хімічним складом вакуолярний сік багатокомпонентний. Він містить вуглеводи, білки, алкалоїди, таніни і т.д. Більшість речовин належить до вторинних продуктів метаболізму, тому вакуоля ще виконує інкреторну (нагромаджувальну) функцію.
1.8 Ергастичні речовини
Ергастичні речовини є продуктами життєдіяльності протопласту. їх можна поділити на запасні (структурні), що тимчасово виключаються з обміну речовин, та екскреторні (рослинні включення), які є вторинними метаболітами і повністю виключені з обміну речовин.
До запасних ергастичних речовин належать запасний крохмаль, ліпідні краплі та білкові включення. Основною запасною формою вуглеводів є крохмаль. Розрізняють крохмаль асиміляційний, що утворюється в хлоропластах, транзитний (транспортний) та запасний або вторинний, що відкладається у вигляді зерен в лейкопластах клітин органів, позбавлених світла (бульби, насіння). На рис. 3.11а показано крохмальні зерна з клітин сім'ядолей
Рис.3.11 а. Запасні поживні речовини в клітинах сім'ядолейгороху (а) та рицитин (б).1- крохмальне зерно, 2 - алейронове зеро.
Білки відкладаються в гіалоплазмі, лейкопластах, вакуолях у вигляді кристалів (кристалоїд) та в аморфному стані (глобоїд) (рис. 3.11 б). Білкові включення ще називають алейроновими зернами.
Ліпіди відкладаються у вигляді ліпідних крапель в гіалоплазмі, особливо в клітинах насіння та плодів. У світловому мікроскопі вони виглядають як сильно заломлюючі світло сферичні тільця.
Екскреторні речовини різноманітні за хімічною будовою. Найбільш розповсюдженими малорозчинні солі щавлевої та оцтової кислот, які відкладаються в вакуолях, утворюючи різноманітні за формою кристали (друзи, рафіди) (рис. 3.13).
3
а б с
Рис. 3.11.б .
Кристалічні включення рослинної клітини .а-клітини луски цибулі, б-клітини кореневища купени, с- клітини черешка бегонії.1 - кристали, 2 - рафіди, 3 - друзи.
Алкалоїди - азотовмісні органічні речовини гетероциклічної природи. На сьогодні їх відомо до 2000, причому більшість має сильні отруйні властивості.
Глікозиди - сполуки цукрів зі спиртами, альдегідами, фенолами. Частина з них відіграє роль пігментів (т.з. антоціани).
Таніни - безазотні похідні фенолів в'яжучого присмаку, які мають протибактеріальні властивості.
Перерахованими хімічними речовинами різноманітність рослинних включень не вичерпується.