

40. Основные углеводы тканей животных, их физиоло-ая роль. Особенности метаболизма фруктозы и галактозы, связь с онтогенезом. Галактоземия, фруктозурия. Углеводы(сахариды)-производные многоатомных спиртов. Делят на группы:олиго-(ди-, трисахариды и т.д. до 10), поли-(гомополисахариды-крахмал, гликоген и гетерополисахариды-целлюлоза, клетчатка), моносахариды(альдозы-пентозы,гексозы и кетозы-фруктоза). В орг-ме явл источником Е для непосредственного использования(глюкоза) или обр-ют депо Е(гликоген), могут синтез-ть соед-я др классов, напр. липиды и некот.АК; явл компонентом ряда сложных соед-ий(нуклеопротеиды, гликопротеины), исп-емых для построения кл.структур. У. животных-глюкоза,играет роль связующего звена м/у энергетическими и пластическими ф-ми, т.к. из глюкозы могут обр-ся все др моносахариды, и наоборот.Метаболизм фруктозы и галактозы вкл пути испол-ия их для синтеза др в-в (гетерополисахаридов, лактозы и др.) и участие в энергообеспечении орг-ма,т.е. фруктоза и галактоза превращаются в печени либо в глюкозу, либо в промежуточные продукты её метаболизма. Т.о., могут быть окислены до СО2 и Н2О или использованы на синтез гликогена и триацилглицеролов. Прич. нарушения метаболизма м.б. дефект ферментов. Метаболизм фруктозы – нач-ся с р-ии фосфорилирования обр-ем фруктозо-6-фосфата при участии ф. гексокиназы и АТФ(р-я ингибируется глюкозой), из-за этого накапливается в крови. Далее ф-6-ф либо превращается в глюкозу ч/з стадии обр-я глюкозо-6-ф и последующим отщеплением фосфорной к-ты, либо подвергается дальнейшим превращениям. Из ф-6-ф под влиянием 6-фосфофруктокиназы и АТФ обр-ся фруктозо-1,6-бисфосфат.(это гл путь включения фруктозы в мышечной ткани, почках, жир.ткани). А в печени есть ф.фруктокиназа, кот катализирует фосфорилирование фруктозы не по 6му, а по 1му атому углерода(эта р-я не блокируется глюкозой). Обр-шийся ф-1-ф расщепляется под действием кетозо-1-фосфатальдолазы на диоксиацетонфосфат и D-глицеральдегид, кот-ый под влиянием триокиназы подвергается фосфорилированию до глицеральдегид-3-фосфата.В этот же промежуточный путь гликолиза переходит и диоксиацетонфосфат. Фруктозурия- врожденный недостаток фруктокиназы, т.е. в орг-ме не обр-ся ф-1-ф. Почечный порог для фруктозы оч низок, поэтому фруктозурия обнар-ся уже при конц-ии фруктозы в крови=0,73 ммоль/л. Галактоза (в основном в печени). Основным источником галактозы явл лактоза пищи, кот в пищ-ом тракте расщепляется до галактозы и глюкозы. Галактозагалактозо-1-фосфат(под действием галактокиназы и АТФ)глюкозо-1-фосфат(гексозо-1-фосфатуридилтрансфераза в присутствии УДФ-глюкозы) и уридиндифосфатгалактоза(УДФ-галактоза). Глюкозо-1-ф либо переходит в глюкозо-6-ф, либо под влиянием фосфотазы обр-ет свободную глюкозу, а УДФ-галактоза (под д-ем УДФ-глюкозы-4-эпимеразы); УДФ-глюкоза+пирофосфат (под д-ем УДФ-глюкоза-пирофосфорилаза); глюкозо-1-ф+УТФ. Галактоземия-стр 32
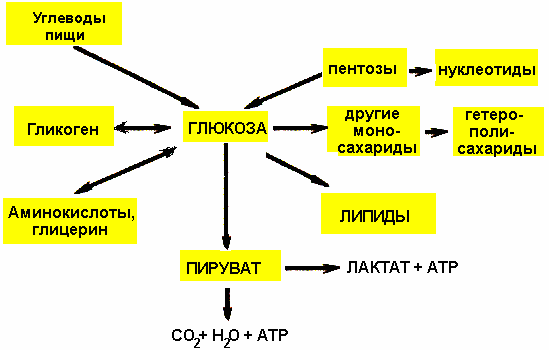
41. Основные углеводы пищи, переваривание и всасывание, возможные нарушения процессов (пр.). Общая схема источников и путей расходования глюкозы в орг-ме.+стр.129.Основные пищевые У.-сахароза, крахмал, целлюлоза, глюкоза. Суточная потребность=400-500г. Глюкоза может обр-ся в орг-ме из АК, из глицерина, входящего в состав жиров(триацилглицеринов). и из лактата. У.пищи в пищ-ом тракте распадаются на мономеры при действии гликозидаз-ферментов, катализирующих гидролиз гликозидных связей.Ротовая полость(рН=6,5-6,8)-кратковременное действие, переваривание крахмала и гликогена под д-ем α-амилазы слюны до декстринов и небольшого кол-ва мальтозы. Желудок (рН=1,5-2,5)-не сод-ит ферментов, расщепляющий сл.углеводы, амилаза инактивируется, но в глубоких слоях пищевого комка амилаза еще есть и полисахариды расщепляются с обр-ем декстринов и мальтозы. 12-перстная кишка(рН= нейтр)-распад крахмала под д-ем α-амилазы поджелуд.сока. Тонкий киш.-амилаза в составе сока поджел.железы расщепляет гликозидные связи,обр-ся олигосахариды и дисахарид мальтоза(амилаза не гидрализует дисахариды);из остатков глюкозы, соед-ых 1,6-гликозидной связью, обр-ся дисахарид изомальтоза.Олиго- и дисахариды гидролизуюся специф-ми гликозидазами тонкого киш-ка.Эти ферменты синтезируются в кл.киш-ка.Продукты полного переваривания У.-моносахариды глюкоза, галактоза и фруктоза-ч/з кл.киш-ка поступают в кровь. При всасывании из киш-ка в кровь моносахариды проникают ч/з кл-ые мембраны путем облегченной диффузии. Глюкоза и галактоза всасываются активным транспортом, кот создается Na,K-ATФазой. Этот мех-м обеспечивает перенос моносахаридов против градиента конц-ии(может функционировать тогда, когда конц-я глюкозы и галактозы в киш-ке<их конц-ии в крови). Нарушения процессов - непереносимость лактазы(приобретенная или временная) при заболеваниях ЖКТ, при некот.инфекциях. Проявление-диарея после приема молока=лактоза поступает в тонкий киш-к, где сбраживается киш-ой флорой с обр-ем газов и к-т; к-ты вследствии осмотического давления привлекают много воды в киш-к , возникает понос. Метеоризм явл.причиной киш-ых колик.Недостаточность лактазы исчезает после излечения болезни.Опасна времен-ая недост.у грудных детей
42. Аэробный гликолиз, последовательность р-ий, связь с общими путями катаболизма. Физиол-ая роль процесса. Аэробный распад(стр.21)-осн. путь катабализма глюкозы у аэробных орг-ов,3 части:1)аэробный гликолиз;2)общий путь катаболизма(окис-е декарбоксилирование пирувата и ЦТК);3)митохондриальная цепь переноса электронов.В результате глюкоза распадается до СО2 и Н2О, а освобождающаяся Е исполь-ся для синтеза АТФ.Распад глюкозы до пирувата делят на 2 этапа:от глюкозы до глицеральдегидфосфата и от него до пирувата.1этап:вкл-е фосфатных остатков в гексозы и превращение гексозы в триозу. Глюкоза(глюкокиназа)использ-е АТФ глюкозо-6-фосфат(фосфоглюкоизомераза) фруктозо-6-фосфат(фосфофруктокиназа)исполь-е АТФфруктозо-1,6-бисфосфат (альдольное расщепление - альдолаза фруктозо-1,6-бисфосфат и фосфотриоизомераза)диоксиацетонфосфат и глицеральдегидфосфат. Диоксиацетонфосфат глицеральдегидфосфат. 2 этап:вкл-ет р-ии, связанные с синтезом АТФ. Глицеральдегидфосфат (дегидрогеназа глицеральдегидфосфата)с обр-ем АТФ1,3-дифосфоглицерат (фосфоглицераткиназа) с обр-ем АТФ3-фосфоглицерат(фосфоглицеромутаза)2-фосфоглицерат (енолаза) фосфоенолпируват (пируваткиназа) с обр-ем АТФпируват. Происходит 6 р-ий дегидрирования-одна на стадии глицеральдегидфосфата(обр-2 АТФ) и 5 в общем пути катаболизма(обр-е 15 АТФ). С вос-ых коферментов водород передается на О2 воздуха ч/з митохондриальную дых-ую цепь.Всего 20 АТФ, но*2=40, и минус 2 АТФ из 1 этапа, получается 38 моль АТФ на 1 моль глюкозы. 10 первых р-ий локализованы в цитозоле, все остальное в митохондриях. В 1ой р-ии 2 этапа обр-ся НАДН, кот не может передать Н на дых.цепь из-за непроницаемости дых.цепи для НАДН. Поэтому перенос Н происходит с помощью челночных мех-ов. Их суть: НАДН в цитозоле вос-ет соед-е, способное проникать в митохондрию; в митох-ии это соед-е окис-ся, восс-я внутримитохонд-ый НАД и вновь переходит в цитозоль(пр.-глицерофосфатный и малат-аспартатный челнок). Физиол-ая роль: 1)происходит во всех тканях и органах(мышцы, почки, сильнее в нервной ткани), но они используют и др. способы синтеза АТФ; 2)более зависим мозг, использует 100г глюкозы и 20% поступающего О2 в сутки; 3)значение в печени-обр-е субстратов и Е для синтеза липидов; 4)активация при длительной мышечной работе и при «2ом дыхании»
43. Анаэробные окис-я глюкозы (анаэробный гликолиз), последовательность р-ий, физиол-ое значение, регуляция. Судьба молочной к-ты. Лактатацидоз. Анаэробный гликолиз=цикл Кори =глюкозолактатный(стр14). В кл. чела есть ф.лактатдегидрогеназа, катализирующее обратное превращение пирувата в молочную к-ту. 10 ф. с аэробного гликолиза совместно с лактатдегидрогеназой обеспечивают синтез АТФ в отсут О2.При этом акцептором Н служит пируват,кот превращается в лактат, выполняющего ф-ю накопителя Н. В анаэробном окис-е,не нуждающемся в митохонд-ой дых.цепи,АТф обр-ся за счет 2х р-ий субстратного фосфорилирования. Обр-ся 4 моль АТФ, и минус 2 АТФ на начальные стадии= 2 АТФ на 1 моль глюкозы. Утилизация лактата в орг-ме: из тканей выводится в кровь и утилизируется в печени, превращаясь в глюкозу, в пр.глюконеогенеза. Физиологическое значение: 1)единственный процесс в орг-ме, продуцирующий Е без О2; 2)быстрый короткий пр.;3)возможен во всех кл. и тканях; 4)наиболее интенсивно протекает в: эритроциты(т.к. нет митохондрий), мышцы, опухолевые кл., эмбриональная ткань(у новорожденных). Значение глюкозы для анаэробной микрофлоры:1)явл основным источником Е; 2)лежит в основе молочнокислого брожения у бактерий, спиртового брожения у дрожжей. Регуляция: эф.Пистера-снижение скорости потребления глюкозы и накопления лактата в присутствии О2. Лактатацидоз- накопление лактата в кл. крови, слюне снижение рНразвитие лактатацидоза. Причины: гипоксия; гиповитаминозы В1, В2, РР; интенсивная мыш.работа. Последствия:болевой синдром в мышцах;развитие утомления.
44. Аэробный катаболизм глюкозы, этапы катаболизма, регуляция, участие вит-ов, физиол-ая роль процесса.Аэробный распад-основной путь катабализма глюкозы у аэробных орг-ов,3 части:1)аэробный гликолиз;2)общий путь катаболизма(окис-е декарбоксилирование пирувата и ЦТК);3)митохондриальная цепь переноса электронов.В результате глюкоза распадается до СО2 и Н2О, а освобождающаяся энергия используется для синтеза АТФ. Окислит-ое декарбок-е пирувата:в результате ацетил-КоА, вос-ный НАД и СО2. 3 фермента- пируватдекарбоксилаза (сод-т коф.тиаминдифосфат, кот явл производным вит.В1 и пирофасфатной к-ты), дегидролипоат-ацетилтрансфераза(коф.липоевая к-та и коф.КоА) и дегидрогеназа дигидролипоевой к-ты(коф.ФАД). 5 коферментов- ФАД(В2), тиаминпирофосфат, липоевая к-та(прочно связаны с ферментами комплекса) и НАД(РР), КоА(нахся в свободно растворенном состоянии и служат акцепторами ацетильного остатка-окис-ся в ЦТК и атомов водорода-с НАДН поступает в цепь переноса электронов). 1 р-я:от пирувата отщепляется СО2, а ацетильный остаток присоед-ся к атому серы липоевой к-ты с обр-ем ацетиллипоат-Е2. 2 р-я: перенос ацетильного остатка на КоА, обр-ся ацетил КоА и дигидролипоевая к-та( в составе ацетилтрансферазы). 3 р-я: акцептором Н служит НАД, обр-е начальной формы-дегидролипоевой к-ты. Пируватдегидрогеназный комплекс-митохондриальный фермент: он соед-н с внутр мембраной со стороны матрикса; пируват поступает к комплексу из матрикса и сюда же освобождаюся ацетил-КоА и НАДН. ЦТК: в цикле лимонной к-ты ацетильный остаток(входящий в ацетил-КоА) обр-ет ряд доноров Н, кот при участии дегидрогеназ поступает в дых.цепь и окис-ся до СО2 и Н2О. Конденсация ацетильного остатка(из ацетил-КоА) и оксалоацетата при участии цитратсинтазы цитрат(аконитаза)изоцитрат(изоцитратдегидрогеназа)α-кетоглутарат(сходен с пируватом). α-кетоглутарат+НS-KoA+НАД(+)Сукцинил-КоА+НАДН2 +СО2. Сукцинил-КоА + ГДФ + Н3РО4 (сукцинаттиокиназа) сукцинат+HS-КоА+ГТФ. Энергия ГТФ может трансформироваться в АТФ при д-ии нуклеозиддифосфаткиназы: ГТФ+АДФГДФ+АТФ.Сукцинат(сукцинатдегидрогеназой)фумарат(фумараза)малат(малатдегидрогеназа)оксалоацетат. ВЫВОД:в общем пути катаболизма распадается пируват с обр-ем 3 молекул СО2(1 при окис-ом декарб-ии пирувата и 2 за счет окис-я ацетильного остатка в ЦТК-р-я 3и 4. Сукцинатдегидрогеназа частью молекулы выступает в матрикс(нах-ся центр связывания сукцината), а частью-погружена во внутр мембрану(центр связывания убихинона). Все остальные ферменты нах-ся в матриксе митохондрий. АТФ=12.Регуляция-скорость р-ий зависит от скорости дыхания митохондрий и окис-ого фосфорилирования. Регенерация НАД и пируватдегидрогеназный комплекс поддерживает скорость обр-я пирувата и ацетил-КоА. Физиолог-ая роль-выработка Е=36.
45. Пентозофосфатный путь катаболизма глюкозы, окислит-ые р-ии, распространение, энергетическая и анаболическая ф-ии.Пентозофосфатный путь(ПФП,фосфоглюконатный,апотомический)-+стр 104. обеспечивает кл гидрированным НАДФ для вос-ых синтезов и пентозами для синтеза нуклеотидов(т.е. выполнение анаболических ф-ий) и НАДФН исполь-ся в мех-ах обезвреживания токсических в-в и в разрушении фагоцитированных МКО. Энергетическая ф-я проявляется в подключении конечных продуктов данного цикла к гликолизу. 2 части: окис-ый путь(глюкозо-6фосат до рибозо-5-фосфат) и неокис-ый путь синтеза пентоз. Коф. дегидрогеназ явл НАДФ, кот вос-ся в НАДФH и исполь-ся кл в р-ии вос-ния и гидроксилирования. ПФП (окис-ый и неокис-ый) поставляет кл пентозофосфаты, необходимые для синтеза НК и коф. (НАД, ФАД, КоА). Все р-ии проходят в цитозоле кл. Р-ии неокис-ого этапа явл обратимыми, поэтому становится возможным синтез гексоз из пентоз. Некот метаболиты неокис-ого пути явл также и метаболитами гликолиза.Зн, что оба процесса тесно связаны и в зависимости от потребностей кл возможны переключения с одного пути на др. При сбалансированной потребности в НАДФН и пентозах в кл происходит окис-ый путь синтеза пентоз. Если потребности в пентозах превышают потребности в НАДФН, то окис-ый путь шунтируется за счет исполь-я метаболитов гликолиза: фруктозо-6-фосфат и глицероальдегидфосфат в р-ях неокис-ого пути превращаются в пентозы. Если НАДФН необходим больше, чем пентозы, то есть 2 варианта:1) при высоком энерг-ом статусе кл излишки пентоз путем обратных р-ий неокис-ого пути превращаются в фруктозо-6-фосфат и глицероальдегидфосфат, из кот в процессе глюконеогенеза обр-ся глюкоза; 2) при низком энерг-ом статусе кл из пентоз обр-ся глицероальдегидфосфат и фруктозо-6-фосфат, кот затем вкл-ся в гликолиз. Значение: активно функционирует в органах, где синтез-ся большие кол-ва липидов-в печени, жир.ткани, молочной железе,коре надпочечников.
46. Глюконеогенез. Роль лактата, АК, глицерина. Значение процесса, регуляция. Роль биотина. Глюконеогенез (синтез глюкозы из пирувата)-стр 55. Гликолиз протекает в цитозоле, а часть р-ий глюконеогенеза происходит в митохондриях. Протекает так же как гликолиз(вопр. № 42), но в обратном направлении. Но 3(II-IV) р-ии гликолиза необратимы, т.е. на этих стадиях р-ии отличны.1 р-я: пируват(пируваткарбоксилаза, его коф. явл биотин)фосфоенолпируват; 2 р-я: фруктозо-1,6-бисфосфатфруктозо-6-фосфат; 3 р-я: глюкозо-6-фосфатглюкоза. Каждая из необратимых р-ий гликолиза вместе с соответствующей ей р-ей глюконеогенеза обр-ет субстратный цикл. Эти циклы служат точками приложения регуляторных мех-ов, направляющих метаболизм на путь или гликолиза, или глюконеогенеза. Синтез глюкозы из АК и глицерина. При катаболизме АК в качестве промеж-ых продуктов обр-ся пируват или оксалоацетат, кот могут вкл-ся в путь глюконеогенеза на стадии 1ого субстратного цикла. Глицерин обр-ся при гидролизе жиров и может превращаться в глюкозу. АК и глицерин исполь-ся для синтеза глюкозыпри голодании или при низком содер-ии углеводов в рационе(углеводное голодание). В этих условиях служит для обеспечения глюкозой мозга, а др ограны обеспечиваются Е за счет окис-я. Глюконеогенез из лактата. Обр-е лактата явл тупиковым путем метаболизма(единственный способ ее исполь-я это превращение в пируват). Из кл, в кот происходит гликолиз, обр-щийся лактат поступает в кровь и улавливается печенью, где и превращается в пируват. В печени частично окис-ся, частично превращается в глюкозу-цикл Кори или глюкозолактатный. Часть пирувата в мышцах путем трансаминирования в аланин, кот транспортируется в печень, и здесь обр-ся пируват-глюкозоаланиновый цикл. В результате цикла Кори работающие мышцы добывают 2 АТФ за счет расходования 6АТФ в печени, и на стадии обр-я глицеральдегидфосфата расходуется НАДН. Значение. Вся глюкоза в орг-ме в конечном счет окис-ся до СО2 и Н2О аэробным путем. Аэробный распад служит вспомогательным путем использования Е глюкозы, имеющее локальное(напр,в эритроцитах) или временное(в работающей мышце) значение. Продукт аэробного распада- лактат- окис-ся аэробным путем. За сутки синтезируется 80 г глюкозы(в основном в печени).Биол-ое значение-возвращение лактата в метаболический фонд углеводов и обеспечение глюкозой мозга при недостатке углеводов(углеводное или полное голодание).Роль биотина(вит.Н) в глюконеогенезе, в процессе кот 16 из 22 АК преобразуются в глюкозу. Этот процесс важен для поддержания стабильного уровня сахара в крови. Т.о. биотин стабилизирует сод-е сахара в крови.
47. Биосинтез и мобилизация гликогена: физиол-ое значение. Зависимость от ритма питания. Регуляция. Гликогенозы и агликогенозы. Гликоген-запасная форма глюкозы, накапливается в кл во время пищеварения и м/у приемами пищи. Осуществляется след-ми ф.: 1) Фосфоглюкомутаза –глюкозо-6-фосфатглюкозо-1-фосфат; 2) Глюкозо-1-фосфат-уридилтрансфераза- синтез УДФ-глюкозы. Необратимость обеспеч-ся гидролизом обр-ся дифосфата; 3) Гликогенсинтаза – обр-ет α1,4-гликозидные связи и удлиняет гликогеновую цепочку;4) Амило-α1,4-α1,6-гликозилтрансфераза,"гликоген-ветвящий" ф – переносит фрагмент в 6 остатков глюкозы на соседнюю цепь с обр-ем α1,6-гликозидной связи.Гликоген печени расщепляется при снижении конц-ии глюкозы в крови,в основном м/у приемами пищи. Ч/з 12-18 ч голодания запасы истощаются. В мышцах кол-во гликогена умень-ся во время физ-ой нагрузки. Гликоген здесь исполь-ся для обеспечения глюкозой работы миоцитов. Т.о., мышцы и печень исполь-ют гликоген только для собственных нужд. Гликогенолиз активируется при недостатке свободной глюкозы в кл, а зн. и в крови (голодание, мышечная работа). При этом уровень глюкозы крови поддерживает только печень, в кот имеется глюкозо-6-фосфатаза, гидролизующая фосфатный эфир глюкозы. Обр-мая в гепатоците свободная глюкоза выходит ч/з плазматическую мембрану в кровь. В гликогенолизе участвуют 3 ф.:1) Фосфорилаза (коф. пиридоксальфосфат) – расщепляет α-1,4-гликозидные связи с обр-ем глюкозо-1-фосфата. Ф. работает до тех пор, пока до точки ветвления (α1,6-связи) не останется 4 остатка глюкозы; 2) α(1,4)-α(1,6)-Глюкантрансфераза – переносит фрагмент из 3х остатков глюкозы на др цепь с обр-ем новой α1,4-гликозидной связи. При этом на прежнем месте остается один остаток глюкозы и "открытая" α1,6-гликозидная связь; 3) Амило-α1,6-глюкозидаза, ("деветвящий" ф.) – гидролизует α1,6-гликозидную связь с высвобождением свободной глюкозы. Обр-ся цепь без ветвлений, вновь служащая субстратом для фосфорилазы. Синтез: почти во всех тканях, но наибольшие запасы нах-ся в печени и скелетных мышцах. Накопление гликогена в мышцах -в период вос-я после работы(особенно при приеме богатой углеводами пищи). В печени гликоген накапливается после еды, при гипергликемии. Такие отличия печени и мышц обусловлены наличием различных изоферментов гексокиназы, фосфорилирующей глюкозу в глюкозо-6-фосфат. Для печени хар-ен изофермент (гексокиназа IV)- глюкокиназа. Отличиями этого ф. от др. гексокиназ явл:1)низкое сродство к глюкозе: захват глюкозы печенью только при ее высокой конц-ии в крови (после еды);2)продукт р-ии (глюкозо-6-фосфат) не ингибирует ф., а в др тканях гексокиназа чувствительна к такому влиянию. Это позволяет гепатоциту захватывать глюкозы больше, чем он может сразу же утилизовать. Благодаря особенностям глюкокиназы гепатоцит эффективно захватывает глюкозу после еды ипосле метаболизирует ее в любом направлении. При норм-ых конц-ях глюкозы в крови ее захват печенью не производится. Регуляция: Скорость гликолиза регулируется потребностями кл в АТФ; регулируется вкл-е глюкозы в процесс гликолиза, она может поступать из 2ух источников: свободная глюкоза и гликоген; Ф., вкл-щие глюкозу в гликолиз, явл регуляторными(напр.гексокиназа).Регуляция гормонов: адреналин и глюкагон активируют распад гликогена печени до глюкозы и вызывают увел-е уровня сахара в крови(синергизм); Глюкокортикоиды, не влияя на тонус мускулатуры сосудов и распад гликогена печени, создают условия, при кот даже небольшие конц-ии адреналина увел-ют артериальное давление и вызывают гипергликемию в результате гликогенолиза в печени(пермиссивное д-е). Гликогенозы-стр 50.
48. Регуляция обмена углеводов и поддержания физиол-го уровня глюкозы в крови. Гипо- и гипергликемии. Уровень глюкозы в крови регулируется гормонами – инсулином, глюкагоном, адреналином, соматотропином и кортизолом. Инсулин снижает уровень глюкозы в крови при её повышении, облегчает проникновение её в кл, способствует отложению глюкозы в тканях в виде гликогена. При снижении уровня глюкозы в крови глюкагон, адреналин, соматропин и кортизол "тормозят" захват глюкозы клетками и обеспечивают трансформацию гликогена в глюкозу. Обмен углеводов регулируется факторами, влияющие на активность ф.: конц-ия субстратов, сод-е метаболитов отдельных р-ий, кислородный режим, проницаемость биол-их мембран, конц-я коф., t и т.д. Регуляция осуществляется при участии ЦНС и гормонов. Гипергликемия-стр 39. Гипогликемия- вызвано понижением ф-ий эндокринных желез. Наблюдают при гипотиреозе, аддисоновой болезни, аденоме островков Лангерганса поджелудочной железы(повышенная секреция инсулина), голодании, продолжительной физ-ой работе, иногда отмечают при беременности и лактации, при введении больших доз инсулина больным диабетом. Сопровождается глюкозурией-из-за сниженного почечного порога для глюкозы.
49. Особенности обмена глюкозы в различных тканях (мышцы, эритроциты, мозг, жировая ткань, печень). Зависимость путей использования глюкоза от ритма и хар-ра питания. Мозг, не имеет запаса гликогена, поэтому он нуждается в постоянном поступлении глюкозы. Углеводы – единственный источник, за счёт кот в норме покрываются энергетические расходы мозга. Ткань мозга поглощает около 70%глюкозы, выделяемой печенью. Инсулин оказывает слабое влияние на поступление и исполь-е глюкозы мозговой тканью. Дело в том, что мембраны кл мозга проницаемы для глюкозы и могут ее исполь-ть без посредничества инсулина. Кл мозга используют только глюкозу, поэтому важно, чтобы уровень глюкозы в крови всегда поддерживался выше критического уровня, что явл одной из важнейших ф-ий с-мы, регулирующей уровень глюкозы в крови.Если уровень глюкозы падает слишком низко (до 20-50 мг/дл), развивается симптоматика гипогликемического шока, кот харак-ся прогрессирующим повыш-ем раздражимости мозга, что ведет к потере сознания, коме. Жир ткань- В условиях больших аэробных нагрузок проникновение глюкозы в жир кл тормозится. Сразу после еды глюкоза попадает в жир ткань и немедленно используется или же откладывается в виде гликогена. Кл в мышечной и жир ткани захватывают глюкозу одним и тем же способом. ГЛЮТ 4 -основной белок, благодаря кот глюкоза проникает в кл орг-ма. Транспортные белки глюкозы находятся в 2х местах:в плазматической мембране и в везикулах, накапливающих ГЛЮТ 4. Когда уровень глюкозы низкий, в жир и мышечной ткани только небольшое кол-во ГЛЮТ 4 удается отыскать на пов-ти плазматической мембраны. Подавляющее ее большинство нах-ся в везикулах. Мембранные транспортеры глюкозы нужны для перенесения глюкозы в кл, даже если ее уровень и низок. Сущ-ет постоянная утилизация ГЛЮТ 4. Мышечная ткань, особенно при активной работе, извлекает из крови значительное кол-во глюкозы. Так же как и в печени, в мышцах из глюкозы синтезируется гликоген. Сущ-ют 2 состояния, когда мышцы потребляют большое кол-во глюкозы. 1)тяжелые физ-ие нагрузки-потребление глюкозы не требует большого кол-ва инсулина, т.к. мышечные волокна становятся более проницаемы для глюкозы даже при отсут инсулина за счет процессов сокр-я. 2) период ч/з несколько ч. после приема пищи - в это время конц-я глюкозы в крови высока и поджелуд. железа секретирует большое кол-во инсулина. Высокая конц-я инсулина обеспечивает быстрый транспорт глюкозы в мышечные кл. Если мышцы после приема пищи не работают, а глюкоза в больших колич-ах еще поступает в кл, она запасается в виде гликогена вместо того, чтобы использоваться в энергетических целях. Гликолиз явл одним из источников Е для мышечного сокр-я. Из продуктов гликолиза – молочной и пировиноградной к-т, в фазе покоя в мышцах, вновь синтезируется гликоген. Эритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохр-е структуры и ф-ии гемоглобина, целостность мембран и обр-е Е для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% - в пентозофосфатном пути(ПФП).Конечный продукт анаэробного гликолиза- лактат выходит в плазму крови и исполь-ся в др кл, прежде всего гепатоцитах. АТФ, обр-ся в анаэробном гликолизе, обеспечивает работу Nа+, К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной р-ях. Особенность анаэробного гликолиза в эритроцитах по сравнению с др кл - присутствие в них ф. бисфосфоглицератмутазы, кот катализирует обр-е 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата. Обр-ся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином.Глюкоза в эритроцитах исполь-ся и в ПФП, окисл-ый этап кот обеспечивает обр-е коф. НАДФН, необходимого для вос-ия глутатиона. Печень- происходит синтез глюкозы (до 250 г в сутки). Глюконеогенез контролируется гормонами. Кортизол, глюкагон и адреналин стимулируют этот процесс, а инсулин, напротив, подавляет. Наиболее важными субстратами явл лактат, поступающий из мышечной ткани и эритроцитов, АК из ЖКТ (глюкогенные АК) и мышц (аланин), а также глицерин из жир тканей. Жир к-ты и др источники ацетил-КоА не могут использоваться в орг-ме для биосинтеза глюкозы, поскольку ацетил-КоА, образ-ся при β-окис-ии в ЦТК, полностью окисляется до СО2, в то время как в глюконеогенезе исходным продуктом явл оксалоцетат.
50. Ненасыщенные и полиненасыщенные (ПНЖК) жирные к-ты. Зависимость их конц-ии от питания. w-3 и w-6 жирные кислоты как предшественники синтеза простагландинов и лейкотриенов. Мононенасыщенные ЖК(с одной двойной связью)-напр. кротоновая(С4), пальмитолеиновая (С16), олеиновая(С18).Полиненасыщенные(с 2 или более двойными связями)-линолевая(С18-с 2мя связями), линоленовая(С18-с 3мя связями), арахидоновая(С20-с 4мя) явл незаменимыми; эти к-ты условно объединены в группу по наз-ем «вит. F».стр 26 в словаре про вит. w-3 и w-6 ПНЖК не синтезируются в орг-ме, поэтому поступают только с пищей. Они принимают участие в физиолог-их и пластических процессах, формируют эйкозаноиды (простагландины, лейкотриены, тромбоксаны), могут эстерифицироваться и гидролизироваться до тканевых глицеролипидов. Но неэстерифицированные w-6 к-ты трансформируются в w-6 эйкозаноиды, кот явл мощными агонистами сигнальных рецепторов, поддерживающих местное и системное воспаление. Лейкотриены - это провоспалительные медиаторы, кот синтезируются из арахидоновой к-ты (полиненасыщенной w-6 к-ты) с участием фермента 5-липооксигеназы. 2ая группа биолог-ки активных в-в – производных арахидоновой к-ты, поддерживающих местное и системное воспаление – это простагландины, синтезируемые с помощью ф. циклооксигеназы.