

2 модуль / Lekcija_8_Ruseckaja_Hormones
.pdfСГМУ им. В.И. Разумовского |
лектор: |
кафедра биохимии |
Русецкая Н.Ю. |
Лекция № 8. Тема: «Гормоны»
План.
1.Понятие о гормонах.
2.Классификация гормонов.
3.Свойства гормонов.
4.Физиологическое действие гормонов.
5.Механизм действия гормонов.
6.Тканевые гормоны
1. Понятие о гормонах.
ГОРМОНЫ (hormao, греч – приводить в движение, побуждать) – группа биологически активных веществ, выделяемых железами внутренней секреции в небольших количествах в кровь и выполняющих функцию регуляторов активности метаболических процессов в клетках.
2. Классификация гормонов.
1.физиологическая: по железам внутренней секреции,
2.биохимическая: по химической структуре гормонов и их растворимости,
3.механизмам действия.
По химической структуре различают:
1.сложные белки (гликопротеины) – фоллитропин (ФСГ), лютропин, тиреотропин (ТТГ).
2.Гормоны пептидной и белковой природы. Это гормоны гипофиза, гипоталамуса, поджелудочной железы, паращитовидных желез.
3.Гормоны, производные аминокислот (фен и тир): гормоны мозгового слоя надпочечников норадреналин, адреналин, дофамин. В своей основе имеют спирт катехол (1,2- диоксибензол), поэтому называются катехоламинами. Это гормоны мозгового вещества надпочечников (адреналин и норадреналин).
4.Гормоны стероидной природы. Это гормоны половых желез и коры надпочечников. Стероидные гормоны синтезируются из холестерина и имеют в основе кольцо стерана (циклопентанпергидрофенантрена):
3. Свойства гормонов.
Гормоны независимо от их природы обладают следующими основными свойствами:
1.дистантный характер действия для большинства гормонов (осуществляют свою функцию на расстоянии),
2.высокая биологическая активность (эффективно действуют на клетки в очень низких концентрациях (около 10-6 - 10-11 моль/л).
3.специфичность проявляется в их действии только на определенные клетки – «клеткимишени».
4.короткий период жизни.
4. Физиологическое действие гормонов.
Гормоны действуют в рамках функциональной иерархии (лестницы), т.е. в ступенчатой последовательности действия. Теснейшая связь нервной и гормональной систем происходит в этой последовательности посредством гипоталамуса, где происходит преобразование нервных импульсов в факторы гуморальной регуляции. В ответ на нервный импульс в гипоталамусе вырабатывается нейрогормон. Изучено 10 гормонов гипоталамуса (рилизинг-факторы). Эти гормоны действуют на орган-мишень – переднюю долю гипофиза. Гормоны передней доли гипофиза поступают в кровь, достигают эндокринной железы. Образование гормонов в эндокринных железах происходит непрерывно. Синтезированные гормоны транспортируются в клетки-мишени либо белками плазмы (стероидные и тиреоидные), либо самостоятельно.
5. Механизм действия гормонов
Гормоны – это факторы регуляции обменных процессов. Большинство гормонов обладает дистантным действием, т.е. вырабатываясь в клетках эндокринных желез, гормоны поступают в кровь и достигают органов, на которые оказывают свое специфическое действие. Биологическое

СГМУ им. В.И. Разумовского |
лектор: |
кафедра биохимии |
Русецкая Н.Ю. |
действие гормонов проявляется через их взаимодействие с рецепторами клеток-мишеней и дальнейшее образование химического сигнала внутри клетки, например, изменение скорости метаболических процессов.
По механизму передачи гормонального сигнала в клетку-мишень гормоны делят на 2 группы:
1.цитозольный (прямой) – стероидные гормоны и тироксин.
2.мембранно-внутриклеточный (непрямой) – гормоны белково-пептидной природы и амины, кроме тироксина.
1.Стероидные гормоны и тироксин растворимы в липидах мембран и проникают в клетку, (далее в клеточное ядро и действуют на геном).
Гормоны стероидной природы проникают в клетку-мишень и связываются со специфическими цитоплазматическими белками-рецепторами, к которым они имеют очень высокое сродство. Образующиеся гормон-рецепторные стероид-белковые комплексы перемещаются в ядро клетки, где образуются комплексы с хроматином – ядерные стероидные комплексы.
Ядерные стероидные комплексы изменяют доступность участков ДНК для считывания информации (транскрипции) и, таким образом, влияют на синтез специфических м-РНК, от которых зависит синтез определенных белковферментов в клетке (м-РНК выходят из ядра в
ЦЗ, связываются с рибосомами, которые синтезируют белки-ферменты).
Таким образом, действие стероидных гормонов осуществляется на уровне генома.
2. Водорастворимые белково-пептидные гормоны и катехоламины. Эти гормоны не проникают через двойной слой мембранных липидов. Рецепторы (по химической природе гликопротеины богатые сиаловыми кислотами) для этих гормонов расположены на внешней поверхности клеточной мембраны. Поэтому белково-пептидные гормоны и катехоламины действуют на клеточные ферменты через посредников:
1.циклический аденозинмонофосфат (ц-АМФ)
2.циклический гуанозинмонофосфат (ц-ГМФ)
3.ионы кальция
4.инозитолтрифосфат (ИФ3)
5.диацилглицерид (ДАГ)
Механизм действия аденилатциклазной системы До момента действия гормона эти компоненты находятся в диссоциированнном состоянии,
а после образования гормон-рецепторного комплекса происходят изменения конформации G- белка. В результате одна из субъединиц G-белка приобретает способность связываться с ГТФ. Комплекс «G-белок-ГТФ» активирует мембранный фермент аденилатциклазу. Аденилатциклаза
начинает активно превращать молекулы АТФ в ц-АМФ.
ц-АМФ обладает способностью активировать особые ферменты - протеинкиназы, которые катализируют реакции фосфорилирования различных белков с участием АТФ (киназы – фосфат-трансферазы – переносят остаток фосфорной кислоты от АТФ на субстрат, в данном случае на белки). При этом в состав белковых молекул включаются остатки фосфорной кислоты. Главным результатом этого процесса фосфорилирования является изменение активности фосфорилированного
белка-фермента (активация фермента путем обратимой ковалентной модификации). В различных типах клеток фосфорилированию в результате активации аденилатциклазной системы подвергаются белки с разной функциональной активностью. Например, это могут быть ферменты,
СГМУ им. В.И. Разумовского |
лектор: |
кафедра биохимии |
Русецкая Н.Ю. |
ядерные белки, мембранные белки. В результате реакции фосфорилирования белки-ферменты могут становиться функционально активными или неактивными. Такие процессы будут приводить к изменениям скорости биохимических процессов в клетке-мишени.
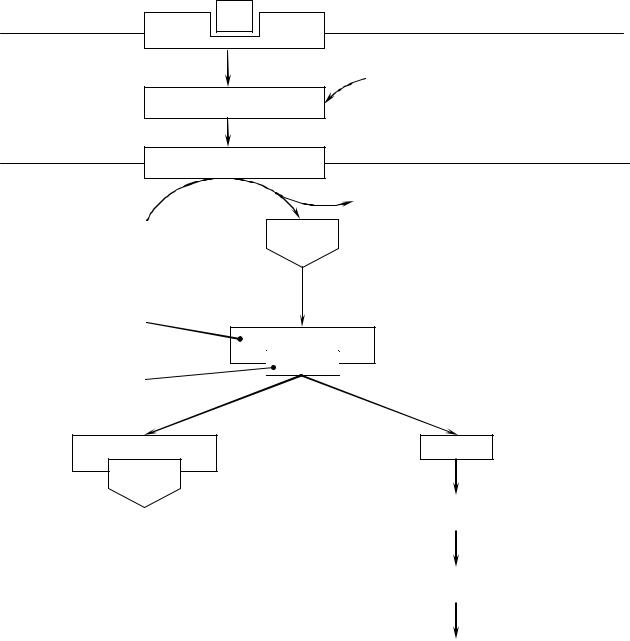
Схема аденилатциклазного механизма действия гормонов
Г
G-белок
Аденилатциклаза
АТФ |
цАМФ |
Регуляторная
субъединица
Каталитическая
субъединица
цАМФ
ГТФ
Мембрана
ФНФН
Неактивная А-протеинкиназа
Активная А-киназа
Фосфорилирование белков и ферментов
Изменение их активности
Клеточный ответ
Активация аденилатциклазной системы длится очень короткое время, потому что G-белок после связывания с аденилатциклазой начинает проявлять ГТФ-азную активность. После гидролиза ГТФ G-белок восстанавливает свою конформацию и перестает активировать аденилатциклазу. В результате прекращается реакция образования ц-АМФ.
Кроме участников аденилатциклазной системы в некоторых клетках-мишенях имеются белки-рецепторы, связанные с G-белками, которые приводят к торможению аденилатциклазы. При этом комплекс “GTP-G-белок” ингибирует аденилатциклазу.
Когда останавливается образование ц-АМФ, реакции фосфорилирования в клетке прекращаются не сразу: пока продолжают существовать молекулы ц-АМФ - будет продолжаться и процесс активации протеинкиназ. Для того, чтобы прекратить действие ц-АМФ, в клетках
СГМУ им. В.И. Разумовского |
лектор: |
кафедра биохимии |
Русецкая Н.Ю. |
существует специальный фермент - фосфодиэстераза, который катализирует реакцию гидролиза 3',5'-ц-АМФ до АМФ.
Примеры гормонов, действующих через ц-АМФ: кортиколиберин, АКТГ, паратгормон, кальцитонин, глюкагон, катехоламины.
Некоторые вещества, обладающие ингибирующим действием на фосфодиэстеразу, (например, алкалоиды кофеин, теофиллин), способствуют сохранению и увеличению концентрации ц-АМФ в клетке. Под действием этих веществ в организме продолжительность активации аденилатциклазной системы становится больше, то есть усиливается возбуждающее действие гормона адреналина. Под действием транквилизаторов (фенозепам) ускоряется разрушение ц-АМФ, прерывается возбуждающее действие адреналина (транквилизаторы применяется при стрессе, в качестве снотворного).
Механизм действия гормонов через ионы Са2+, инозитолтрифосфат (ИФ3) и диацилглицерин (ДАГ)
Кроме аденилатциклазной или гуанилатциклазной систем существует также механизм передачи информации внутри клетки-мишени с участием ионов кальция, диацилглицерида (ДАГ) и инозитолтрифосфата (ИФ3).
Диацилглицерид и инозитолтрифосфат - это вещества, которые являются продуктами гидролиза сложного липида - фосфатидилинозита. ИФ3 образуется в результате действия специального фермента - фосфолипазы “С”, который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора.
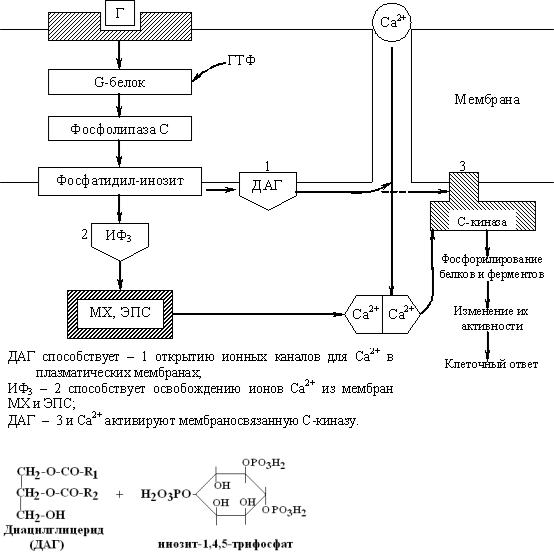
Схема непрямого механизма действия гормонов через ионы Са2+, инозитолтрифосфат (ИФ3) и диацилглицерин (ДАГ)
Фосфолипаза «С» гидролизует фосфоэфирную связь в молекуле фосфатидил- инозитол-4,5-бисфосфата и в результате образуются диацилглицерин (ДАГ) и инозитолтрифосфат (ИФ3).
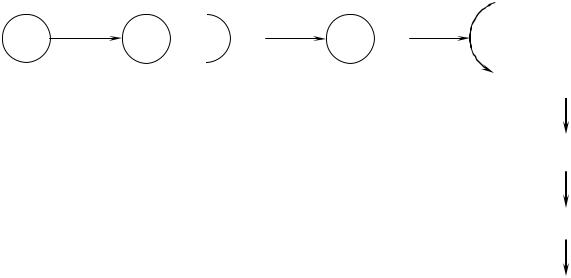
СГМУ им. В.И. Разумовского |
лектор: |
кафедра биохимии |
Русецкая Н.Ю. |
Известно, что образование диацилглицерина (ДАГ) и инозитолтрифосфата (ИФ3) приводит к освобождению из мембран МХ и ЭПС ионов кальция. Концентрация ионизированного кальция внутри клетки увеличивается. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое приводит к физиологическому ответу клетки на действие гормона.
В работе этого механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок - кальмодулин. Это низкомолекулярный белок (17 кДа), на 30% состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са+2. Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са+2 происходят конформационные изменения молекулы кальмодулина и комплекс “Са+2-кальмодулин” становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты - аденилатциклазу, фосфодиэстеразу, Са+2,Мg+2- АТФазу и различные протеинкиназы.
В разных клетках при воздействии комплекса “Са+2-кальмодулин” на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других - ингибирование реакции образования ц-АМФ.
Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са+2- кальмодулин будет отличаться.
Примеры гормонов, действующих через ИФ3 и ДАГ: гонадотропин, вазопрессин, катехоламины, медиатор ацетилхолин, рилизинг-факторы (тиреолиберин).
Схема активации внутриклеточных ферментов ионами Са2+
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Неактивная |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Са2+-КМ-киназа |
|
Са2+ |
|
|
Са2+ + |
КМ |
Са2+ КМ |
|||||
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
Активная |
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
Са2+-КМ-киназа |
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
Фосфорилирование |
||
|
мембрана |
|
|
|
|
белков и ферментов |
||||
|
Изменение их |
|
активности |
Кальцийсвязывающий – 1 белок |
|
цитоплазмы – кальмодулин |
Клеточный ответ |
|
Механизм действия инсулина Действие инсулина начинается с его связывания со специфическим рецептором на поверхности
клетки-мишени. Рецептор инсулина – тирозиновая протеинкиназа, фосфорилирующая белки по ОН-группам тирозина. Рецептор состоит из двух α– и двух β-субъединиц - гликопротеинов с углеводной частью на поверхности мембраны. В отсутствие гормона инсулиновые рецепторы не проявляют тирозинкиназной активности. Присоединение инсулина к центру связывания на α– субъединицах активирует фермент и происходит фосфорилирование β-субъединиц по нескольким тирозиновым остаткам. В результате этого активная тирозиновая протеинкиназа начинает
СГМУ им. В.И. Разумовского |
лектор: |
кафедра биохимии |
Русецкая Н.Ю. |
фосфорилировать другие внутриклеточные белки и ферменты, что приводит к изменению их активности.
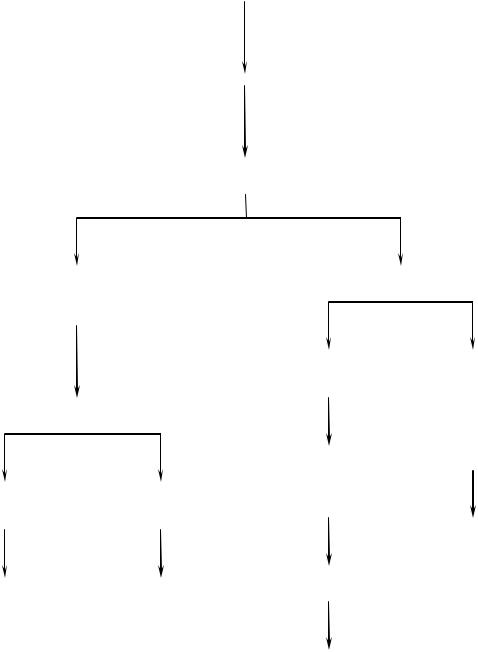
Схема механизма действия инсулина
Связывание инсулина с рецептором на клеточной
Аутофосфорилирование
Поглощение гормонрецепторного комплекса путем
Фосфорилирование |
Активация фосфолипазы и |
|||
клеточных белков- |
фосфатидил-инозитолкиназы |
|||
субстратов инсулинового |
|
|
|
|
|
|
Фосфорилирова |
Активация и |
|
|
|
ние фосфатидил- |
перемещение белков- |
|
Каскад реакций активации |
инозитов |
|
транспортеров |
|
|
|
глюкозы к |
||
|
|
|
|
|
специфических протеинкиназ |
|
|
плазматическим |
|
|
|
Мобилизация Са |
2+ |
мембранам жировых |
|
|
|
|
|
|
|
и активация |
|
и мышечных клеток |
Фосфорилирование |
Фосфорилирование |
специфических |
|
|
цитозольных белков |
белков транскрипции |
протеинкиназ |
|
|
и ферментов |
в ядре |
|
|
Увеличение их |
|
|
|
|
проницаемости для |
|
|
Фосфорилирование |
глюкозы |
|
Изменение их |
Индукция либо |
|
||
белков и ферментов |
|
|||
активности |
репрессия синтеза |
|
белков и ферментов |
||
|
Изменение их
активности
Инсулин активирует ферменты фосфолипазу и фосфатидилинозитолкиназы, следствием чего является образование ИФ3 (механизм действия ИФ3 см. выше) и перемещение белковтранспортеров глюкозы в плазматическую мембрану клеток жировой и мышечной ткани, что ускоряет трансмембранный перенос глюкозы в эти клетки.
Снижение чувствительности клеток к инсулину опосредуется двумя механизмами:
1. Комплекс рецептор-инсулин захватывается внутрь клетки эндоцитозом с последующим разрушением части рецепторов в лизосомах, а часть возвращается в плазматическую мембрану.
СГМУ им. В.И. Разумовского |
лектор: |
кафедра биохимии |
Русецкая Н.Ю. |
2. Фосфорилирование инсулинового рецептора по остаткам серина и треонина снижает его сродство к инсулину.
Роль инсулина в регуляции обмена веществ:
1.индуцирует синтез ферментов гликолиза, пентозного цикла и гликогенсинтазу.
2.тормозит липолиз в адипоцитах, активируя фосфодиэстеразу и уменьшая внутриклеточную концентрацию цАМФ.
3.активирует ферменты синтеза жирных кислот и ТАГ, ацетил-КоА-карбоксилазу и глицеролфосфатацилтрансферазу.
6. Тканевые гормоны
Ктканевым гормонам относят биологически активные вещества, проявляющие свое регулирующее действие на обменные процессы, вырабатывающиеся клетками определенных тканей и действующими либо местно (в данной ткани), либо дистантно.
Ктканевым гормонам и гормоноподобным веществам относятся:
1.Биогенные амины – серотонин, ацетилхолин, гистамин, ГАМК.
2.Простагландины (тромбоксаны, лейкотриены образуются из арахидоновой кислоты).
3.Эндорфины.
4.Гормоны ЖКТ (гастрин, секретин).
5.Цитокины.
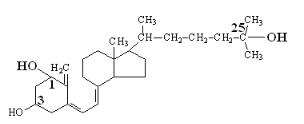
Кальцитриол (стероидный гормон).
- это активная форма витамина Д3 – холекальциферол. В печени холекальциферол под действием специальной гидроксилазы превращается в кальцидиол (25-оксихолекальциферол), который в почках под действием специальной гидроксилазы превращается в Кальцитриол – 1,25диоксихолекальциферол. Специальные гидроксилазы активируются под действием паратгормона.
кальцитриол Органы-мишени: тонкий кишечник (увеличение всасывания кальция и фосфата), кости
(мобилизация кальция и фосфата). Кальцитриол взаимодействует в ядрах клеток с хроматином, стимулируя процесс синтеза белков.
Гиповитаминоз Д – рахит у детей, у взрослых – остеомаляция.