

Документ Microsoft Word (3)
.docxПоксвирусы представляют собой крупные сложноорганизованные вирусы с необычной морфологией. Генетический материал вируса связан с белками в центральной дисковидной структуре, известной как нуклеоид. Нуклеоид окружён мембраной и двумя боковыми телами неизвестной функции. Вирус имеет наружную оболочку с большим количеством белков на её поверхности. Весь вирион слегка плейоморфен (то есть способен изменять форму и размер в зависимости от условий) и может принимать форму от овальной до блоковидной[80]. Мимивирус является одним из крупнейших описанных вирусов и имеет икосаэдрический капсид диаметром 400—500 нм. Белковые филаменты, отходящие от поверхности вириона, достигают 100 нм в длину[81][82]. В 2011 году исследователи обнаружили ещё более крупный вирус на океаническом дне вблизи побережья Чили. Вирус, которому было дано временное название Megavirus chilensis, можно увидеть даже в обычный оптический микроскоп[83].
Геном[править | править вики-текст]
|
Генетическое разнообразие у вирусов |
|
|
Свойства |
Параметры |
|
Нуклеиновая кислота |
|
|
Форма |
|
|
Количество цепей |
|
|
Полярность |
|

Реассортимент может стать причиной появления нового высокопатогенного штамма человеческого гриппа
Вирусы демонстрируют огромное количество вариантов организации генома; в этом смысле они более разнообразны, чем растения, животные, археи и бактерии. Существуют миллионы различных типов вирусов[6], но только примерно 5000 из них были детально описаны[5]. Генетический материал вируса может быть представлен либо ДНК, либо РНК, соответственно, вирусы подразделяют на ДНК-содержащие и РНК-содержащие. Подавляющее большинство вирусов являются РНК-содержащими. Вирусы растений чаще всего содержат одноцепочечную РНК, а бактериофаги, как правило, обладают двухцепочечными ДНК[84].
Вирусный геном может быть кольцевым, как у полиомавирусов, или линейным, как у аденовирусов. Форма генома не зависит от типа нуклеиновой кислоты. У многих РНК-содержащих вирусов и некоторых ДНК-содержащих вирусов геном часто представлен несколькими молекулами (частями), в связи с чем он называется сегментированным. У РНК-содержащих вирусов каждый сегмент часто кодирует только один белок, и обычно эти сегменты упаковываются в один капсид. Однако присутствие всех сегментов не всегда обязательно для инфекционности вируса, как это демонстрируют вирус мозаики костра и некоторые другие вирусы растений[66].
Вирусные геномы независимо от типа нуклеиновый кислоты практически всегда бывают либо одноцепочечным, либо двухцепочечным. Двухцепочечный геном включает пару комплементарных цепей нуклеиновой кислоты, а одноцепочечный — только одну цепь. Геном вирусов некоторых семейств (например, Hepadnaviridae) частично одноцепочечный и частично двуцепочечный[84].
Для большинства РНК-содержащих вирусов и некоторых вирусов с одноцепочечной ДНК определяют полярностьнуклеиновой кислоты в зависимости от того, комплементарна ли она вирусной мРНК. Молекула РНК с положительной полярностью (плюс-цепь) имеет ту же последовательность нуклеотидов, что и мРНК, поэтому, по крайней мере, какая-то её часть может незамедлительно начать транслироваться клеткой-хозяином. РНК с отрицательной полярностью (минус-цепь) комплементарна мРНК, поэтому до начала трансляции на ней должна быть синтезирована положительная РНК при помощи фермента РНК-зависимой-РНК-полимеразы. Названия цепей ДНК для вирусов, содержащих одноцепочечную ДНК, сходны с таковыми для РНК: кодирующая цепь комплементарна мРНК (-), а некодирующая является её копией (+)[84]. Однако геномы нескольких типов ДНК- и РНК-содержащих вирусов представлены молекулами, имеющими различную полярность, то есть транскрипции может подвергаться любая цепь. Таковы, например, геминивирусы — вирусы растений, содержащие одноцепочечную ДНК, — и аренавирусы — вирусы животных с одноцепочечной РНК[85].
Размер генома широко варьирует у различных видов. Самым маленьким одноцепочечным ДНК-геномом обладает цирковирус из семейства Circoviridae: его геном кодирует лишь два белка и содержит всего 2000 нуклеотидов. Один из самых крупных геномов обнаружен у мимивируса: он содержит свыше 1,2 млн. пар оснований и кодирует более тысячи белков[86]. Как правило, РНК-содержащие вирусы имеют меньший геном, чем ДНК-содержащие — размер их генома ограничен из-за большей вероятности ошибок во время репликации[38]. При большем размере генома ошибки, произошедшие во время его репликации, сделали бы вирус нежизнеспособным или неконкурентоспособным. Чтобы преодолеть это ограничение, РНК-вирусы часто имеют сегментированный геном — это уменьшает вероятность того, что ошибка в одном из сегментов окажется фатальной для всего генома. Напротив, ДНК-содержащие вирусы обычно имеют более крупные геномы благодаря большей точности их репликативных ферментов[87]. Однако вирусы, содержащие одноцепочечные ДНК, являются исключением из этого правила — скорость накопления мутаций в их геномах приближается к таковой для вирусов, содержащих одноцепочечные РНК[88].
Генетические изменения происходят у вирусов по разным механизмам. В их число входят случайные замены отдельных оснований в РНК или ДНК. В большинстве случаев эти точечные мутации являются «молчащими» — они не изменяют структуру белков, кодируемых мутантными генами, но иногда в результате таких изменений вирус может приобрести эволюционные преимущества, такие как устойчивость к противовирусным препаратам[89]. Дрейф антигенов имеет место тогда, когда в геноме вируса происходят масштабные изменения. Это может быть результатом рекомбинации или реассортимента. Когда это случается с вирусом гриппа, результатом может статьпандемия[90]. РНК-вирусы часто существуют как квазивиды или смесь вирусов одного вида, но с чуть различающимися нуклеотидными последовательностями генома. Такие квазивиды являются главной мишенью для естественного отбора[91].
Сегментированный геном даёт эволюционные преимущества: различные штаммы вируса с сегментированным геномом могут обмениваться генами и производить потомков с уникальными характеристиками. Это явление называется реассортиментом[92].
Генетическая рекомбинация — это процесс внесения разрыва в молекулу нуклеиновой кислоты с последующим «сшиванием» её с другими молекулами нуклеиновой кислоты. Рекомбинация может происходить между геномами двух вирусов, когда они заражают клетку одновременно. Исследования эволюции вирусов показали, что у изученных видов рекомбинация широко распространена[93]. Рекомбинация характерна как для РНК-, так и для ДНК-содержащих вирусов[94][95].
Жизненный цикл[править | править вики-текст]
Основная статья: Жизненный цикл вируса
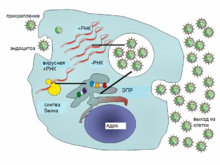
Типичный жизненный цикл вируса на примере вируса гепатита C
Вирусы не размножаются клеточным делением, поскольку не имеют клеточного строения. Вместо этого они используют ресурсы клетки-хозяина для образования множественных копий самих себя, и их сборка происходит внутри клетки.
Условно жизненный цикл вируса можно разбить на несколько взаимоперекрывающихся этапов (обычно выделяют 6 этапов[96]):
-
Прикрепление представляет собой образование специфичной связи между белками вирусного капсида ирецепторами на поверхности клетки-хозяина. Это специфичное связывание определяет круг хозяев вируса. Например, ВИЧ поражает только определённый тип человеческих лейкоцитов. Это связано с тем, что оболочечныйгликопротеин вируса gp120 специфично связывается с молекулой CD4 — хемокиновым рецептором, который обычно встречается на поверхности CD4-положительных T-лимфоцитов. Этот механизм обеспечивает инфицирование вирусом только тех клеток, которые способны осуществить его репликацию. Связывание с рецептором может вызватьконформационные изменения белка оболочки (или белка капсида в случае безоболочечного вируса), что в свою очередь служит сигналом к слиянию вирусной и клеточной мембран и проникновению вируса в клетку.
-
Проникновение в клетку. На следующем этапе вирусу необходимо доставить внутрь клетки свой генетический материал. Некоторые вирусы также переносят внутрь клетки собственные белки, необходимые для её реализации (особенно это характерно для вирусов, содержащих негативные РНК). Различные вирусы для проникновения в клетку используют разные стратегии: например, пикорнавирусы впрыскивают свою РНК через плазматическую мембрану, а вирионы ортомиксовирусов захватываются клеткой в ходе эндоцитоза и попадают в кислую среду лизосом, где происходит депротеинизация вирусной частицы, после чего РНК в комплексе с вирусными белками преодолевает лизосомальную мембрану и попадает вцитоплазму. Вирусы также различают по тому, где происходит их репликация: часть вирусов (например, те же пикорнавирусы) размножается в цитоплазме клетки, а часть (например, ортомиксовирусы) в её ядре. Процесс инфицирования вирусами клеток грибов и растений отличается от инфицирования клеток животных. Растения имеют прочную клеточную стенку, состоящую из целлюлозы, а грибы — из хитина, так что большинство вирусов могут проникнуть в них только после повреждения клеточной стенки[97]. Однако почти все вирусы растений (включая вирус табачной мозаики) могут перемещаться из клетки в клетку в форме одноцепочечных нуклеопротеиновых комплексов через плазмодесмы[98]. Бактерии, как и растения, имеют крепкую клеточную стенку, которую вирусу, чтобы попасть внутрь, приходится повредить. Но в связи с тем, что клеточная стенка бактерий намного тоньше таковой у растений, некоторые вирусы выработали механизм впрыскивания генома в бактериальную клетку через толщу клеточной стенки, при котором капсид остаётся снаружи[99].
-
Лишение оболочек представляет собой процесс потери капсида. Это достигается при помощи вирусных ферментов или ферментов клетки-хозяина, а может быть и результатом простой диссоциации. В конечном счёте вирусная геномная нуклеиновая кислота освобождается.
-
Репликация вирусов подразумевает, прежде всего, репликацию генома. Репликация вируса включает синтез мРНК ранних генов вируса (с исключениями для вирусов, содержащих положительную РНК), синтез вирусных белков, возможно, сборку сложных белков и репликацию вирусного генома, которая запускается после активации ранних или регуляторных генов. Вслед за этим может последовать (у комплексных вирусов с крупными геномами) ещё один или несколько кругов дополнительного синтеза мРНК: «поздняя» экспрессия генов приводит к синтезу структурных или вирионныхбелков.

Отпочковывание вируса от клетки-хозяина
-
Вслед за этим происходит сборка вирусных частиц, позже происходят некоторые модификации белков. У вирусов, таких как ВИЧ, такая модификация (иногда называемая созреванием) происходит после выхода вируса из клетки-хозяина[100].
-
Выход из клетки. Вирусы могут покинуть клетку после лизиса, процесса, в ходе которого клетка погибает из-за разрыва мембраны и клеточной стенки, если такая есть. Эта особенность есть у многих бактериальных и некоторых животных вирусов. Некоторые вирусы подвергаются лизогенному циклу, где вирусный геном включается путём генетической рекомбинации в специальное место хромосомы клетки-хозяйки. Тогда вирусный геном называется провирусом, или, в случае бактериофага, профагом[101]. Когда клетка делится, вирусный геном также удваивается. В пределах клетки вирус в основном не проявляет себя; однако в некоторый момент провирус или профаг может вызвать активацию вируса, который может вызвать лизис клеток-хозяев[102].
Активно размножающийся вирус не всегда убивает клетку-хозяина. Оболочечные вирусы, в том числе ВИЧ, обычно отделяются от клетки путём отпочковывания. В ходе этого процесса вирус обзаводится своей оболочкой, которая представляет собой модифицированный фрагмент клеточной мембраны хозяина или другой внутренней мембраны[103]. Таким образом, клетка может продолжать жить и продуцировать вирус.
Особенности жизненного цикла различных групп[править | править вики-текст]
Генетический материал внутри вирусных частиц и способ его репликации, значительно отличается у различных вирусов.
-
ДНК-содержащие вирусы. Репликация генома у большинства ДНК-содержащих вирусов происходит в клеточном ядре. Если клетка имеет соответствующий рецептор на своей поверхности, эти вирусы проникают в клетку либо путём непосредственного слияния с клеточной мембраной (напр. герпесвирусы), либо — что бывает чаще — путём рецептор-зависимого эндоцитоза. Большинство ДНК-содержащих вирусов полностью полагаются на синтетический аппарат клетки-хозяина для производства их ДНК и РНК, а также последующего процессинга РНК. Однако вирусы с крупными геномами (например, поксвирусы) могут сами кодировать большую часть необходимых для этого белков. Геному вируса эукариот необходимо преодолеть ядерную мембрану для того, чтобы получить доступ к ферментам, синтезирующим ДНК и РНК, в случае же бактерифагов ему достаточно просто проникнуть в клетку[104][105].
-
РНК-содержащие вирусы. Репликация таких вирусов обычно происходит в цитоплазме. РНК-содержащие вирусы можно подразделить на 4 группы в зависимости от способа их репликации. Механизм репликации определяется тем, является ли геном вируса одноцепочечным или двухцепочечным, вторым важным фактором в случае одноцепочечного генома является его полярность (может ли он непосредственно служить матрицей для синтеза белка рибосомами). Все РНК-вирусы используют собственную РНК-репликазу для копирования своих геномов[106].
-
Вирусы, использующие обратную транскрипцию. Эти вирусы содержат одноцепочечную РНК (Retroviridae, Metaviridae, Pseudoviridae) или двухцепочечную ДНК (Caulimoviridae и Hepadnaviridae). РНК-содержащие вирусы, способные к обратной транскрипции (ретровирусы, например, ВИЧ), используют ДНК-копию генома как промежуточную молекулу при репликации, а содержащие ДНК (параретровирусы, например, вирус гепатита B) — РНК[107]. В обоих случаях используется обратная транскриптаза, или РНК-зависимая-ДНК-полимераза. Ретровирусы встраивают ДНК, образующуюся в процессе обратной транскрипции, в геном хозяина, такое состояние вируса называется провирусом. Параретровирусы же этого не делают, хотя встроенные копии их генома могут давать начало инфекционным вирусам, особенно у растений[108]. Вирусы, использующие обратную транскрипцию, восприимчивы к противововирусным препаратам, ингибирующим обратную транскриптазу, в том числе к зидовудину и ламивудину.
Действие на клетки[править | править вики-текст]

Микрофотография, показывающая цитопатические эффекты, вызванные вирусом простого герпеса первого типа. Тест Папаниколау
Диапазон структурных и биохимических эффектов, оказываемых вирусом на инфицированную клетку, очень широк[109]. Они называются цитопатическими эффектами[110]. Большинство вирусных инфекций приводят к гибели клеток-хозяев. Причинами гибели могут быть лизис клетки, изменения клеточной мембраны и апоптоз[111]. Часто причиной гибели клетки является подавление её нормальной активности белками вируса, не все из которых входят в состав вирусной частицы[112].
Некоторые вирусы не вызывают никаких видимых изменений в поражённой клетке. Клетки, в которых вирус находится в латентном состоянии и неактивен, имеют мало признаков инфекции и нормально функционируют[113]. Это является причиной хронических инфекций, и вирус при них может никак себя не проявлять многие месяцы или годы. Так часто бывает, например, с вирусом герпеса[114][115]. Некоторые вирусы, например вирус Эпштейна-Барр, могут вызывать быстрое размножение клеток без появления злокачественности[116], в то время как другие, такие какпапилломавирусы, могут вызвать рак[117].
Диапазон хозяев[править | править вики-текст]
Вирусы, несомненно, являются самыми многочисленными биологическими объектами на Земле, и по этому показателю они превосходят все организмы, вместе взятые[118]. Они поражают все формы клеточных организмов, включая животных, растения, бактерии и грибы[5]. Тем не менее различные типы вирусов могут поражать только ограниченный круг хозяев, многие вирусы видоспецифичны. Некоторые, как, например, вирус оспы, могут поражать только один вид — людей[119], в подобных случаях говорят, что вирус имеет узкий диапазон хозяев. Напротив, вирус бешенства может поражать различные видымлекопитающих, то есть он имеет широкий диапазон хозяев[120]. Вирусы растений безвредны для животных, а большинство вирусов животных безопасны для человека[121]. Диапазон хозяев некоторых бактериофагов ограничивается одним штаммом бактерий, и они могут использоваться для определения штаммов, вызывающих вспышки инфекционных заболеваний методом фагового типирования[122].
Классификация[править | править вики-текст]
В таксономии живой природы вирусы выделяют в отдельный таксон, образующий в классификации Systema Naturae 2000 вместе с доменами Bacteria,Archaea и Eukaryota корневой таксон Biota[123]. В течение XX века в систематике выдвигались предложения о создании выделенного таксона для неклеточных форм жизни (Aphanobionta Novak, 1930[124]; надцарство Acytota Jeffrey, 1971[125]; Acellularia[126]), однако такие предложения не были кодифицированы[127].
Главной задачей классификации является описание разнообразия вирусов и группировка их на основании общих свойств. В 1962 году Андре Львов,Роберт Хорн и Пауль Турнье были первыми, кто разработал основные принципы классификации вирусов на основании Линнеевской иерархической системы[128]. Основными таксонами в этой системе являются отдел, класс, порядок, семейство, род и вид. Вирусы были разделены на группы по общим свойствам (но не таковым у их хозяев) и типу нуклеиновых кислот в геномах[129]. Позднее был создан Международный комитет по таксономии вирусов. Однако в таксономии вирусов не применяются понятия «отдел» и «царство», поскольку их малый размер генома и высокая частота мутаций затрудняет выяснение родства групп старше порядка. По существу, классификация вирусов по Балтимору является дополнением более традиционной классификации.
Систематику и таксономию вирусов в настоящий момент кодифицирует и поддерживает Международный Комитет по Таксономии Вирусов (International Committee on Taxonomy of Viruses, ICTV), поддерживающий также и таксономическую базу The Universal Virus Database ICTVdB.
Классификация ICTV[править | править вики-текст]
Международный комитет по таксономии вирусов разработал современную классификацию вирусов и выделил основные свойства вирусов, имеющие больший вес для классификации с сохранением единообразия семейств.
Была разработана объединённая таксономия (универсальная система для классификации вирусов). Седьмой отчёт ICTV закрепил для первых пор понятие о виде вируса как о низшем таксоне в иерархии вирусов[130][комм. 3]. Однако к настоящему моменту была изучена лишь небольшая часть от общего разнообразия вирусов, анализ образцов вирусов из человеческого организма выявил, что около 20 % последовательностей вирусных нуклеиновых кислот ещё не было рассмотрено ранее, а образцы из окружающей среды, например, морской воды и океанского дна, показали, что подавляющее большинство последовательностей являются совершенно новыми[131].
Основными таксономическими единицами являются[132]:
Отряд (-virales)
Семейство (-viridae)
Подсемейство (-virinae)
Род (-virus)
Вид (-virus)
Современная классификация ICTV (2012) включает 7 отрядов вирусов: Caudovirales, Herpesvirales, Ligamenvirales, Mononegavirales, Nidovirales,Picornavirales и Tymovirales[133]. Существование восьмого порядка (Megavirales[134]) пока ещё было только предположено. Классификация не выделяетподвиды, штаммы и изоляты. Всего насчитывается 6 порядков, 87 семейств, 19 подсемейств, 349 родов, около 2284 видов и свыше 3000 ещё неклассифицированных вирусов[135][136][137].
Классификация по Балтимору[править | править вики-текст]
Основная статья: Классификация вирусов по Балтимору
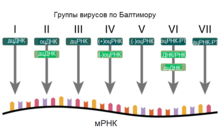
Группы вирусов по Балтимору. Условные обозначения: оц — одноцепочечная, дц — двуцепочечная, РТ — ретровирус или ретроидный вирус.
Лауреат Нобелевской премии биолог Дейвид Балтимор разработал классификацию вирусов по Балтимору[34][138]. Классификация ICTV в настоящее время объединяется с классификацией по Балтимору, составляя современную систему классификации вирусов[139][140].
Классификация вирусов по Балтимору основывается на механизме образования мРНК. Вирусы должны синтезировать мРНК из собственных геномов для образования белков и репликации своей нуклеиновой кислоты, однако каждое семейство вирусов имеет собственный механизм осуществления этого. Вирусные геномы могут быть одноцепоченые (оц) или двухцепочечные (дц), ДНК- или РНК-содержащие, могут использовать или не использовать обратную транскриптазу. Кроме того, одноцепочечные РНК-вирусы могут иметь положительную (+) или отрицательную (-) цепь РНК в составе своего генома.
Эта система включает в себя семь основных групп[141][142]:
-
(I) Вирусы, содержащие двуцепочечную ДНК и не имеющие РНК-стадии (например, герпесвирусы, поксвирусы,паповавирусы, мимивирус).
-
(II) Вирусы, содержащие одноцепочечную молекулу ДНК (например, парвовирусы). В этом случае ДНК всегда положительной полярности.
-
(III) Вирусы, содержащие двуцепочечную РНК (например, ротавирусы).
-
(IV) Вирусы, содержащие одноцепочечную молекулу РНК положительной полярности (например, пикорнавирусы, флавивирусы).
-
(V) Вирусы, содержащие одноцепочечную молекулу РНК негативной или двойной полярности (например, ортомиксовирусы, филовирусы).
-
(VI) Вирусы, содержащие одноцепочечную положительную молекулу РНК и имеющие в своем жизненном цикле стадию синтеза ДНК на матрице РНК, ретровирусы (например, ВИЧ).
-
(VII) Вирусы, содержащие двуцепочечную ДНК и имеющие в своём жизненном цикле стадию синтеза ДНК на матрице РНК, ретроидные вирусы (например, вирус гепатита B)[143].
Дальнейшее деление производится на основе таких признаков как структура генома (наличие сегментов, кольцевая или линейная молекула), генетическое сходство с другими вирусами, наличие липидной оболочки, таксономическая принадлежность организма-хозяина и так далее.
Роль в заболеваниях человека[править | править вики-текст]
Основная статья: Вирусные заболевания

Основные вирусные инфекциичеловека и их возбудители
Примерами наиболее известных вирусных заболеваний человека могут служить простуда (она может иметь и бактериальную этиологию), грипп, ветряная оспа и простой герпес. Многие серьёзные болезни, например,геморрагическая лихорадка Эбола, СПИД, птичий грипп и тяжёлый острый респираторный синдром вызываются вирусами. Относительная способность вируса вызывать заболевание характеризуется термином вирулентность. Некоторые заболевания исследуются на наличие вирусов в числе вызывающих агентов, например, возможна связь между человеческим герпесвирусом 6 типа и нейрологическими заболеваниями, такими как рассеянный склероз исиндром хронической усталости[144]. Идут споры по поводу того, что борнавирус, ранее считавшийся возбудителем нейрологических заболеваний у лошадей, возможно, вызывает психические расстройства у людей[145].
Вирусы имеют различные механизмы, вызывающие болезнь у хозяина, и эти механизмы сильно зависят от вида. Такой механизм на клеточном уровне включает, прежде всего, лизис клеток, приводящий к их смерти. Умногоклеточных организмов, при гибели большого числа клеток, начинает страдать организм в целом. Хотя вирусы подрывают нормальный гомеостаз, приводя к заболеванию, они могут существовать внутри организма и относительно безвредно. В качестве примера можно привести способность вируса простого герпеса первого типа пребывать в состоянии покоя внутри тела человека. Такое состояние называется латентностью[146]. Оно характерно для вирусов герпеса, в том числе вируса Эпштейна-Барр, вызывающегоинфекционный мононуклеоз, а также вируса, вызывающего ветрянку и опоясывающий лишай. Большинство людей переболели по крайней мере одним из этих типов вируса герпеса[147]. Однако такие латентные вирусы могут и принести пользу, поскольку присутствие этих вирусов может вызвать иммунный ответ против бактериальных патогенов, например, чумной палочки (Yersinia pestis)[148].
Некоторые вирусы могут вызывать пожизненные или хронические инфекции, при которых вирус продолжает размножаться в теле организма, несмотря на его защитные механизмы[149]. Так происходит, например, при инфекциях, вызванных вирусами гепатита B и C. Хронически больные люди известны как носители, поскольку они выступают в роли резервуара для заразного вируса[150]. Если в популяции имеется высокая доля носителей, то в этом случае говорят об эпидемии[151].