

Биохимия._Углеводы
.pdfтивируются процессы свободно-радикального окисления. Обычная клетка, и эритроцит в том числе, довольно легко справляются с дополнительной нагрузкой. При недостаточности описываемого фермента перекись водорода накапливается в эритро-ците, усиливается повреждение его мембран и гемолиз.
ГОМЕОСТАЗ ГЛЮКОЗЫ В КРОВИ
Концентрация глюкозы в крови изменяется под влиянием многих гормонов. Ос-
новными гормонами являются глюкагон, адреналин, глюкокортикоиды, сомато-
тропный гормон с одной стороны, и инсулин с другой. Инсулин является единственным гормоном организма, действие которого нацелено на снижение уровня глю-козы крови. Все остальные гормоны увеличивают его.
Уменьшение инсулином концентрации глюкозы в крови достигается следующими путями:
•стимуляция белков-транспортеров на цитоплазматической мембране,
•повышение синтеза глюкокиназы – фермента, получившего название "ловушка для глюкозы",
•активация гликоген-синтазы -и стимуляция ее синтеза, что позволяет превратить излишки глюкозы в гликоген,
•индукция синтеза глюкозо-6-фосфат-дегидрогеназы и 6-фосфоглюконат- дегидрогеназы,
•стимуляция синтеза ферментов гликолиза – фосфофруктокиназы, пируват-киназы, что позволяет вовлечь глюкозу в окислительные процессы.
•вовлечение глюкозы в синтез триацилглицеринов (см Синтез триацилглице-ролов ).
Многие ткани нечувствительны к действию инсулина, их называют инсулиннезависимыми. К ним относятся нервная ткань, стекловидное тело, хрусталик, сетчат-ка, клубочковые клетки почек, эндотелиоциты, семенники и эритроциты.
Глюкагон, адреналин и глюкокортикоиды повышают содержание глюкозы кро-
ви, активируя мобилизациию гликогена (гликогенфосфорилазу), стимулируя синтез ферментов глюконеогенеза (пируваткарбоксилазу, фосфоенолпируваткарбоксикиназу, фруктозо-1,6-дифосфатазу и глюкозо-6-фосфатазу). Глюкокортикоиды, кроме этого, препятствуют проникновению глюкозы в клетку.
ГИПЕРГЛИКЕМИЧЕСКИЕ СОСТОЯНИЯ
Гипергликемическим является состоянием, при котором концентрация глюкозы в крови более 6 ммоль/л.

По происхождению выделяют две группы таких состояний:
1.Физиологические
•алиментарные – связаны с приемом пищи и продолжаются в норме не бо-лее 2 часов после еды.
•нейрогенные – нервное напряжение. Стимулирующее секрецию адреналина и мобилизацию гликогена в печени,
•гипергликемия беременных – связана с относительной недостаточностью инсулина при увеличении массы тела и потребностью плода в глюкозе.
2.Патологические
•Припри заболеваниях гипофиза, коры и мозгового слоя надпочечников, щитовидной железы, при органических поражениях ЦНС и поджелудочной железы.
САХАРНЫЙ ДИАБЕТ
Сахарный диабет (СД) – полиэтиологическое заболевание, связанное:
•со снижением количества β-клеток островков Лангерганса,
•с нарушениями на уровне синтеза инсулина,
•с мутациями, приводящими к молекулярному дефекту гормона,
•со снижением числа рецепторов к инсулину и их аффинности в клеткахмишенях,
•с нарушениями внутриклеточной передачи гормонального сигнала. Выделяют два типа сахарного диабета:
1.Инсулинзависимый сахарный диабет (ИЗСД)- диабет детей и подростков (ювенильный), его доля составляет около 20% от всех случаев СД;
2.Инсулиннезависимый сахарный диабет (ИНЗСД) – диабет взрослых, его доля
–около 80%;
Подразделение типов СД на взрослый и ювенильный не всегда корректно, так как встречаются случаи развития ИНЗСД в раннем возрасте, также ИНЗСД может переходить в инсулинзависимую форму
3. Диабет беременных
Более подробно рассмотрим 1 и 2 типы СД. Развитие ИЗСД обусловлено не-
достаточным синтезом инсулина в
β-клетках островков Лангерганса поджелудочной железы. Среди причин этого в настоящее время на первый план выдвигаются аутоиммунные поражения и инфицирование β-тропными вирусами (вирусы Коксаки, Эпштейна-Бар, эпидемического паротита).
Дополнение
Имеется опасность использования коровьего молока или молочных смесей для вскармливания младенцев из-за возможного развития иммунного ответа на молочный альбумин и переключения иммунной атаки на β-клетки поджелудоч-ной железы.

Для диабета взрослых ведущей причи-
ной является инсулинорезистентность,
возникающая из-за функциональных или структурных нарушений инсулиновых рецепторов.
Сравнительная характеристика типов сахарного диабета
|
ИЗСД |
ИНЗСД |
|
|
|
|
|
Возраст |
Дети, подростки |
Средний, пожилой |
|
|
|
|
|
Начало |
Острое (несколько дней) |
Постепенное (годы) |
|
|
|
|
|
Внешний вид (до лечения) |
Худощавое |
У 40% ожирение |
|
|
|
|
|
Снижение массы тела (до |
Обычно есть |
Не характерно |
|
лечения) |
|||
|
|
||
Концентрация инсулина в |
Снижена в 2-10 раз |
В норме или повышена |
|
крови |
|||
|
|
||
Концентрация С-пептида |
Резко снижена |
В норме или повышена |
|
или отсутствует |
|||
|
|
||
Семейный анамнез |
Отягощен редко |
Часто отягощен |
|
|
|
|
|
Зависимость от инсулина |
Полная |
Только у 20% |
|
|
|
|
|
Склонность к кетоацидозу |
Есть |
Нет |
|
|
|
|
Диагностика.
Диагноз инсулинзависимого сахарного диабета ставится если
1.Имеются классические симптомы (полиурия, полидипсия, снижение массы тела) и концентрация глюкозы натощак в нескольких повторных анализах капиллярной крови более 6,1 ммоль/л.
2.Отсутствие соответствующих симптомов при концентрации глюкозы натощак в нескольких повторных анализах капиллярной крови более 6,1 ммоль/л.
3.В сомнительных (и только!) случаях – отсутствие симптомов в сочетании неоднозначностью результатов анализов – рекомендуется нагрузочная проба с глюкозой. Она заключается в приеме испытуемым глюкозы из расчета 1,5-2,0 г на кг массы тела. Пробы крови отбирают непосредственно перед приемом глюкозы (нулевая ми-
нута, "тощаковый" уровень) и далее через 30, 60, 90 и 120 минут, при необходимости на 180
минуте.

В норме повышение концентрации глюкозы составляет 50-75% к 60 минуте исследования и снижается до исходных величин к 90-120 минутам. В абсолютных единицах по реко-
мендации ВОЗ подъем уровня глю-козы должен быть не более 7,5 ммоль/л при исходном 4,0-5,0 ммоль/л.
Дополнение
Иногда пробы берут только на 0 и 120 минутах, однако это нежела-
тельно, так как упускается до-полнительная информация о состоянии организма. Так, по крутизне восходя-
щей части кривой можно судить об активности n. vagus, отвечающего за секрецию инсулина, о всасывающей функции кишечника, о способности печени усваивать глюкозу. К примеру, "голодная" печень с истощенными запасами гликогена, более активно потребляет глюкозу из крови воротной вены по сравнению с
"сытой", и подъем кривой более плавный. Аналогичная кривая на-блюдается при ухудшении всасывания глюкозы вследствие заболевания слизистой кишечника. При циррозе печени отмечается обратная картина.
Довольно часто у взрослых вместо глюкозной нагрузки используется обычный завтрак, и кровь отбирают через 1, 2 или 2,5 часа после него. Если уровень глюкозы в указанное время не возвращается к норме, то подтверждается ди-агноз сахарного диабета.
Гипергликемические кривые
проявляются повышенным в 2-3 раза уровнем глюкозы крови после нагрузки, что свидетельствует о нарушении гормональных взаимо-
действий. Нормализация показате-лей происходит крайне медленно и завершается не ранее 150-180 минут . Наиболее частой причиной та-ких кривых является скрытый сахарный диабет 1 и 2 типа и повреждении паренхимы печени. Избыток катехоламинов при феохромоцитоме и трийодтиронина при гиперфункции щитовидной железы,
гиперкортицизм, заболевания гипоталамуса и гипофиза также проявляются в виде гипергликемической кривой.
Дополнение
При измерении уровня глюкозы после еды у больных с хорошо контролируе-мым сахарным диабетом результаты должны укладываться в диапазон 7,6- 9,0 ммоль/л. Величины большие 9,0 ммоль/л означают, что дозировка инсулина неправильна или диабет не компенсирован.

Гипогликемические кривые –
повышение концентрации глюкозы не более чем на 25% с быстрым возвращением к исходным значениям. Наблюдаются при аденоме островков Лангерганса, гипотирео-зе, гипофункции коры надпочечников, заболеваниях кишечника и дисбактериозе.
Дополнение
ЭФФЕКТЫ ИНСУЛИНА
Очень быстрые эффекты (секунды)
1.Гиперполяризация мембран чувствительных к инсулину клеток;
2.Активация na+/h+-обменника, что выхывает выход ионов н+, вход в клетку ио-
нов na+;
3.Активация na+/к+-обменника, что выхывает выход ионов na+, вход в клетку ио-нов
k+;
4.Угнетение ca2+-насоса, приводит к задержке ионов са2+ в клетке;
5.Стимуляция транспорта глюкозы в клетку – появление на мембране перенос-чиков
глюкозы;
Быстрые эффекты (минуты)
1.Стимулирование протеинфосфатаз приводит к активации гликогенсинтазы, пируватдегидрогеназы, ГМГ-SКоА-редуктазы, ацетил-S-КоА-карбоксилазы;
2.Увеличивает липогенез за счет создания благоприятного "биохимического климата":
•активирует глюкозо-6-фосфатдегидрогеназу и, что вызывает наработку НАДФН,
•глюкокиназу, что ведет к синтезу ацетил-SКоА,
•ацетил-S-КоА-карбоксилазу и синтазу жирных кислот, повышая синтез жир-ных кислот.
3.Активирует цАМФ-фосфодиэстеразу, снижая тем самым ее активирующее
влияние на протеинкиназу А и реакции катаболизма.
Медленные эффекты (минуты-часы)
1.Активация синтеза глюкокиназы, АТФ-цитрат-лиазы, ацетил-S-КоА-карбо- ксилазы, синтазы жирных кислот, пируваткиназы, люкозо-6-фосфатдегидрогеназы, цитолозольной малатдегидрогеназы.;
2.Увеличение синтеза тРНК для увеличения скорости транскрипции. Однако на-работку мРНК антагонистических ферментов снижает (например, для ФЕПкарбоксикиназы);
3.Повышает фосфорилирование по серину рибосомального белка S6, что сти-мулирует
синтез белка.
Очень медленные эффекты (часы-сутки)
1.Повышает синтез соматомедина, зависимый от гормона роста;
2.Увеличивает рост и пролиферацию клеток, действуя при этом синергично с соматомедином;
3.Стимулирует тирозиновые киназы., вызывает переход клетки из G1 в S-фазу
клеточного цикла.
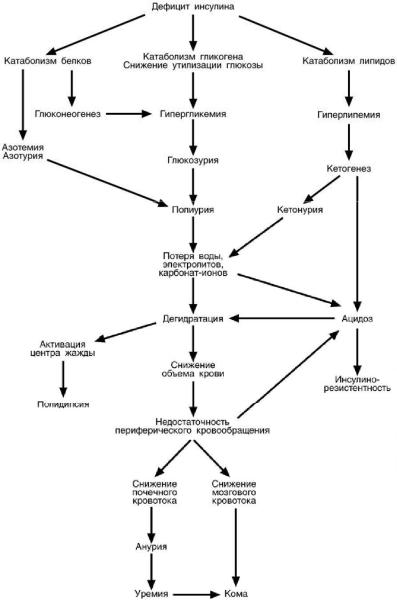
П ОСЛЕДСТВИЯ ДЕФИЦИТА ИНСУЛИНА
Быстрые последствия
1.Гипергликемия – так как отсутствует влияние инсулина и превалирует влияние глюкагона, адреналина, кортизола, гормона роста.
2.Глюкозурия – почечный порог для глюкозы, т.е. концентрация глюкозы в крови при которой она появляется в моче, примерно равен 10,0 ммоль/л. В норме в моче уровень глюкозы 0,8 ммоль/л и до 2,78 ммоль/сут, в других единицах около 0,5 г/сут, при СД количество теряемой глюкозы составляет до 100 г/сут и более.
3.Преобладание катаболизма белков над анаболизмом ведет к накоплению продуктов азотистого обмена, в первую очередь мочевины и ее повышенному выведению. Избыток аминокислот уходит в глю-
конеогенез.
4.Глюкоза и мочевина
осмотически удерживают воду в просвете поччного канальца и возникает полиурия. Объем мочи возрастает в 2-3 раза.
5.Повышенный распад ТАГ в жировой ткани и печени обуславливает аномально высокое окисление жирных кислот и накопле-ние их недоокисленных продуктов – кетоновых тел. Это приводит к кетонемии,
кетонурии и кетоацидозу.
При диабете концентрация кетоновых тел возрастает
в100-200 раз и достигает
350 мг% (норма 2 мг% или
0,1-0,6 ммоль/л).
6.При полиурии с мочой, кроме воды, теряются соли, в частности карбонаты, имеющие щелочной характер. Это усугубляет ацидоз.
7.В результате
.п.п.4,5,6 возникает дегид-

ратация (в тяжелых случаях до 5 л) организма, которая заключается в падении объема крови, обезвоживанию клеток и их сморщиванию (дряблая кожа, запавшие глаза, мягкие глазные яблоки, сухость слизистых), уменьшению артериального давле-ния. Ацидоз вызывает одышку (дыхание Kussmaul, быстрое и глубокое) и дополнительную дегидратацию.
8.Активируется центр жажды и начинается полидипсия.
9.Дегидратация неминуемо приводит к недостаточности кровообращения в тканях – активируется анаэробный гликолиз, накапливается лактат и в дополнение к кетоацидозу возникает лактацидоз.
10.Закисление среды вызывает изменение взаимодействия инсулина с рецепторами, клетки становятся нечувствительными к инсулину – инсулинорезистент-
ность.
11.Ацидоз крови уменьшает концентрацию 2,3-дифосфоглицерата в эритроцитах. Это, повышая сродство гемоглобина к кислороду, создает тканевую гипоксию и усугубляет лактацидоз
Отдаленные последствия
Гипергликемия резко повышает потребление глюкозы инсулиннезависимыми тканями (клетки артериальных стенок, эндотелия, клетки Шванна, эритроциты, хрусталик и сетчатка глаза, семенники и гломерулярные клетки почек), в них вынужден-но
активируется особые пути метаболизма глюкозы. Их интенсивность определяется только доступностью глюкозы:
1.Превращение глюкозы в
сорбитол.
Сорбитол плохо проникает через клеточные мембраны, его накопление в цитозоле приводит
космотическому набуханию клеток и нарушению их функций. Например, возникновение катаракты хрусталика и нейропатий (нарушение осязания) в клетках Шванна
2.Неферментативное гликозилирование различных бел-
ков, изменение их свойств и активация их синтеза за счет избытка энергии:
•увеличивается синтез гликопротеинов базальной мембраны почечных клу-бочков. Это приводит к окклюзии капилляров и нарушению фильтрации
•увеличивается синтез гликопротеинов в сетчатке глаза, что вызывает отек сетчатки и кровоизлияния
•увеличивается синтез гликопротеинов в стекловидном теле
•увеличивается синтез тканевых белков за счет доступности глюкозы и энергии
•гликозилированные белки хрусталика объединяются в крупные агрегаты, рассеивающие свет. Это вызывает помутнение хрусталика и катаракту.
•гликозилирование гемоглобина в эритроцитах, образование HbA1C
•белков свертывающей системы, что увеличивает вязкость крови
•белков ЛПНП, что уменьшает их связывание с рецепторами и повышает концентрацию ХС в крови
•белков ЛПВП, что усиливает их сродство к рецепторам и быструю элиминацию из кровотока

Из-за двух последних нарушений возникают макроангиопатии развивается атеросклероз сосудов мозга, сердца, почек, конечностей. Характерно в основном для ИНЗСД.
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ
При калорийности рациона 2000–3000 ккал суточное потребление углеводов составляет 300-450г. С пищей поступает крахмал, сахароза, лактоза, пищевые волокна (клетчатка и др.). Переваривание углеводов начинается в ротовой полости при участии α- амилазы слюны, которая расщепляет в крахмале α-1,4-гликозидные связи. Полное расщепление крахмала здесь не происходит, так как пребывание пищи во рту кратковременно. Из крахмала в ротовой полости образуются крупные фрагменты – декстрины. Желудочный сок не содержит ферментов, расщепляющих углеводы. Дальнейшее переваривание углеводов происходит в тонком кишечнике. Фермент поджелудочной железы α-амилаза расщепляет α-1,4-гликозидные связи крахмала и декстринов, α-1,6-гликозидные связи расщепляются ферментом кишечного сока – амило-1,6-гликозидазой. При действии двух ферментов образуется дисахарид мальтоза. Амилаза поджелудочной железы не расщепляет β-1.4-гликозидные связи, которыми соединены остатки глюкозы в молекуле целлюлозы. Поэтому пищевые волокна не перевариваются, но они должны присутствовать в рационе, так как улучшают перистальтику, ускоряют чувство насыщения и снижают уровень холестерина в крови, поскольку на них происходит адсорбция желчных кислот и выведение их из организма. Пищевые волокна обязательно должны присутствовать в рационе при ожирении, запорах, атеросклерозе, сахарном диабете. Мальтоза, образовавшаяся из крахмала, а также дисахариды пищи – сахароза и лактоза перевариваются ферментами тонкого кишечника – дисахаридазами. Эти ферменты работают не в просвете кишечника, а на поверхности эпителиальных клеток кишечника. Мальтоза расщепляется мальтазой до 2 молекул глюкозы, лактоза – лактазой до глюкозы и галактозы, сахароза – сахаразой до глюкозы и фруктозы (рис.3). Все моносахариды всасываются, сначала путем облегченной
диффузии, а затем активным транспортом в симпорте с ионами Nа+.
Рис.3. Катаболизм дисахаридов и патогенез дисахаридозов
В крови воротной вены содержатся три моносахарида: глюкоза, фруктоза и галактоза. Все они попадают в печень, где происходит унификация фруктозы и галактозы, т.е. они превращаются в глюкозу – единственный моносахарид, используемый всеми клетками нашего организма.
Дисахаридозы – нарушение переваривания дисахаридов, связанные с недостаточной активностью дисахаридаз. Недостаточная активность ферментов может быть врожденной и

приобретенной. Симптомы врожденных форм проявляются достаточно рано, например, после первого кормления грудным молоком (при дефиците лактазы) или при добавлении в рацион сахара или крахмала. Приобретенные формы могут наблюдаться при заболеваниях кишечника. Нерасщепленные дисахариды вызывают осмотическую диарею, сбраживаются микрофлорой кишечника с образованием углекислого газа, что приводит к метеоризму, коликам.
ОБМЕН ГЛИКОГЕНА
Многие ткани в качестве резервной формы глюкозы синтезируют гликоген. Синтез и распад гликогена обеспечивают постоянство концентрации глюкозы в крови. Синтезгликогена происходит в покое и сытости, как любой анаболический процесс требует энергии. Депонируется гликоген главным образом в печени и мышцах. Глюкоза, поступившая в клетку, фосфорилируется при участии гексокиназы за счет АТФ, при этом образуется глюкозо-6-фосфат, который в ходе обратимой реакции под действием фосфоглюкомутазы превращается в глюкозо-1-фосфат. Затем при участии УТФ глюкозо-1- фосфат превращается в УДФ-глюкозу. Эта молекула используется как донор остатков глюкозы при синтезе гликогена.
Так как гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путем удлинения уже имеющейся молекулы полисахарида, называемой «затравка». К «затравке» последовательно присоединяются остатки глюкозы из УДФглюкозы α- 1,4-гликозидной связью при участии фермента гликогенсинтазы. Разветвленная структура гликогена образуется при участии «фермента ветвления» (рис.4). Регуляторными ферментами в синтезе гликогена являются гликогенсинтаза и гексокиназа. Синтез гликогена увеличивается под влиянием инсулина, а тормозится глюкагоном, катехоламинами, глюкокортикостероидами.
Рис.4. Обмен гликогена печени
Распад гликогена происходит путем последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата. Гликозидная связь расщепляется с присоединением неорганического фосфата, поэтому процесс называется фосфоролизом, а фермент – фосфорилазой. Образовавшийся глюкозо-1-фосфат затем изомеризуется фосфоглюкомутазой до глюкозо-6-фосфата. В печени (но не в мышцах) глюкозо-6-фосфат может гидролизоваться с образованием глюкозы, которая выделяется в кровь. Эту реакцию катализирует глюкозо-6- фосфатаза. Мышечный гликоген не используется для поддержания уровня глюкозы в крови, так как в мышцах нет фермента глюкозо-6-фосфатазы и образование свободной глюкозы там
невозможно, а глюкозо-6-фосфат не может проникать через мембрану клеток. Таким образом, печень запасает глюкозу в виде гликогена не столько для собственных нужд, сколько для поддержания постоянной концентрации глюкозы в крови. Функция мышечного гликогена заключается в освобождении глюкозо-6-фосфата, потребляемого в самой мышце для окисления и использования энергии.
Регуляторными ферментами распада гликогена являются фосфорилаза и глюкозо- 6-фосфатаза. Процесс распада усиливают катехоламины, глюкагон, глюкокортикостероиды; тормозит инсулин.
РАСПАД ГЛЮКОЗЫ В АЭРОБНЫХ И АНАЭРОБНЫХ УСЛОВИЯХ
Катаболизм глюкозы – основной поставщик энергии для большинства клеток организма.
Аэробный распад протекает в условиях достаточного снабжения тканей кислородом. Он включает несколько стадий:
1. Аэробный распад глюкозы до двух молекул пирувата в гиалоплазме клеток,который можно разделить на 2 этапа:
а)подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз.
б) основной этап, сопряженный с синтезом АТФ. В результате серии реакций фосфотриозы превращаются в пируват.
•Превращение пирувата в ацетил-КоА и окисление последнего в цикле Кребса
•Дыхательная цепь
Глюкозо-6-фосфат, образованный в результате фосфорилирования глюкозы с участием АТФ, в ходе следующей реакции превращается в фруктозо-6-фосфат. Это обратимая реакция изомеризации протекает при участии глюкозофосфатизомеразы. Затем следует еще одна реакция фосфорилирования за счет АТФ, катализирует ее фосфофруктокиназа. В ходе этой реакции фруктозо-6-фосфат превращается в фруктозо-1,6-бисфосфат. Этот метаболит далее расщепляется на 2 триозофосфата: фосфоглицериноый альдегид (ФГА) и диоксиацетонфосфат (ДОАФ), реакцию катализирует альдолаза. В последующих реакциях гликолиза используется только ФГА, поэтому ДОАФ превращается с участием фермента триозофосфатизомеразы в ФГА. На этом подготовительный этап заканчивается.
Основной этап аэробного распада включает реакции, связанные с синтезом АТФ. Сначала происходит окисление ФГАпри участии ФГА-дегидрогеназы. Этот фермент является
НАД+-зависимымым. Восстановленный НАД+ затем окисляется в дыхательной цепи (перенос НАДН в митохондрии, где находится дыхательная цепь, происходит при участии челночных механизмов, это связано с тем, что митохондриальная мембрана непроницаема для НАДН). Кроме того, свободная реакция окисления этой реакции концентрируется в макроэргической связи продукта реакции. Продуктом реакции является 1,3-
бисфосфоглицерат. Для фосфорилирования используется Н3РО4. В следующей реакции высокоэнергетический фосфат передается на АДФ с образованием АТФ. Катализирует реакцию фосфоглицераткиназа. АТФ в данной реакции образуется путем субстратного фосфорилирования. Продуктом реакции является 3-фосфоглицерат. В следующей реакции происходит перенос фосфата из 3 положения во второе с образованием 2-фосфоглицерата. Из него при отщеплении воды образуется фосфоенолпируват – макроэргическое соединение, фосфатная группа которого в следующей реакции переносится на АДФ с образованием АТФ при участии пируваткиназы (это вторая реакция субстратного фосфорилирования). Продуктом реакции является пируват.
Во второй стадии пируват превращается в ацетил-КоА, который сгорает в цикле Кребса и дыхательной цепи (третья стадия) до СО2 и Н2О.
Всего при аэробном окислении глюкозы выделяется 32 АТФ.
Анаэробный гликолиз представляет собой распад глюкозы в анаэробных условиях (условиях недостаточного снабжения кислородом), поэтому он не зависит от работы митохондриальной дыхательной цепи. АТФ в гликолизе образуется путем