

Энергия в клетке 201
Глава 11. Энергетический обмен
11.1Энергия в клетке
Впростой прокариотической клетке кишечной палочки E. coli происходит более 1000 химических реакций, которые в сумме составляют клеточный метаболизм. Это количество может показаться чересчур большим, однако при детальном рассмотрении становится ясно, что многие из них имеют общий характер или черты. Фактически, примерно 100 молекул играют ключевую роль в жизни всех клеток на нашей планете. И несмотря на то, что количество реакций просто огромно, видов реакций не так уж и много, а их механизмы часто довольно просты. «Энергетической валютой» для всех форм жизни служит аденозинтрифосфат (АТФ) — рибонуклеотид, содержащий 3 остатка фосфорной кислоты, связанных макроэргическими связями. Разрыв этих связей приводит к выделению большого количества энергии, которая используется в анаболизме, то есть АТФ выступает в роли донора энергии в клетках.
Живые организмы нуждаются в притоке свободной энергии для покрытия трёх основных задач:
1.Выполнение механической работы: сокращение мышц и движение клеток;
2.Активный транспорт ионов и молекул;
3.Синтез макромолекул и других биомолекул из простых предшественников.
Фотосинтезирующие микроорганизмы (фототрофы) используют солнечный свет в качестве источника этой энергии. Хемотрофы (например, животные) получают эту энергию, окисляя те соединения, которые синтезируются фототрофами.
Помимо АТФ в роли источников энергии в живых клетках могут выступать и другие соединения:
1.Нуклеозидтрифосфаты (ГТФ, УТФ и ЦТФ);
2.Фосфоенолпируват (участвует в субстратном фосфорилировании АДФ в АТФ);
3.Креатинфосфат (используется в мышцах для регенерации АДФ в АТФ);
4.1,3-Бисфосфоглицерат (участвует в субстратном фосфорилировании АДФ в АТФ).
202Глава 11 Энергетический обмен
11.2Дыхательная цепь митохондрий
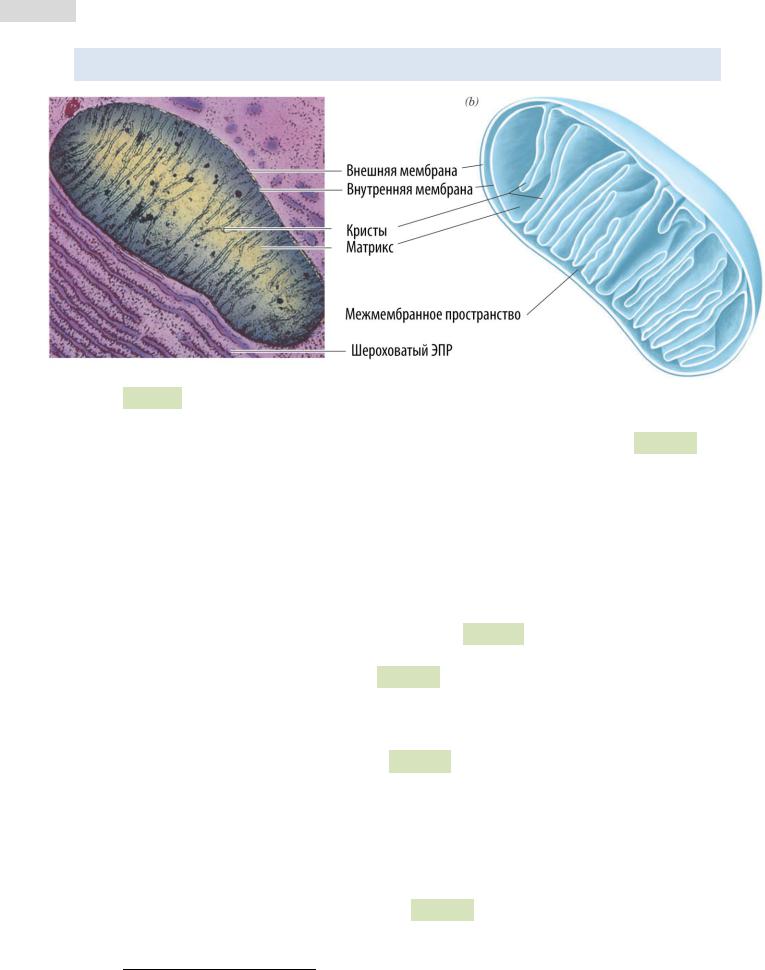
Рис. 118. Митохондрия.
Митохондрии представляют собой двумембранные органеллы (см. Рис. 118 ▲). Внешняя мембрана имеет поры, пропускающие в межмембранное пространство молекулы до 10 кДа (вплоть до небольших белков). Внутренняя мембрана митохондрий очень селективна и содержит множество транспортных систем, переносящих важные соединения из межмембранного пространства в матрикс митохондрий (т.е. во внутреннюю полость).
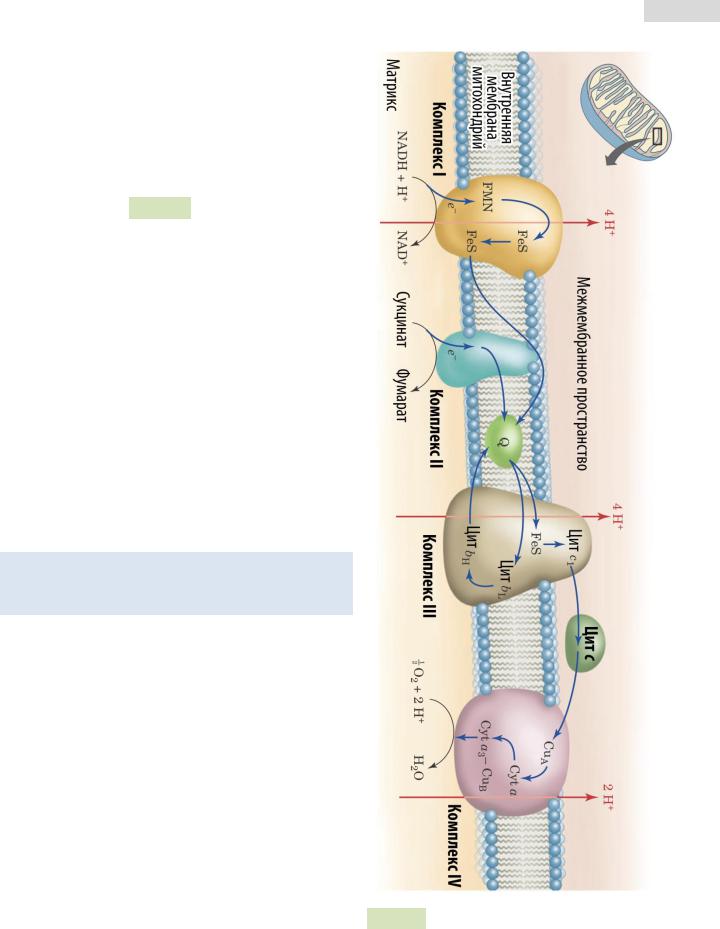
Во внутреннюю мембрану митохондрий встроены белковые комплексы — компоненты дыхательной цепи (цепи переноса электронов или электронтранспортной цепи). Дыхательная цепь митохондрий крайне важна, поскольку отвечает за синтез АТФ в клетке. Компоненты дыхательной цепи (см. Рис. 119 ▼):
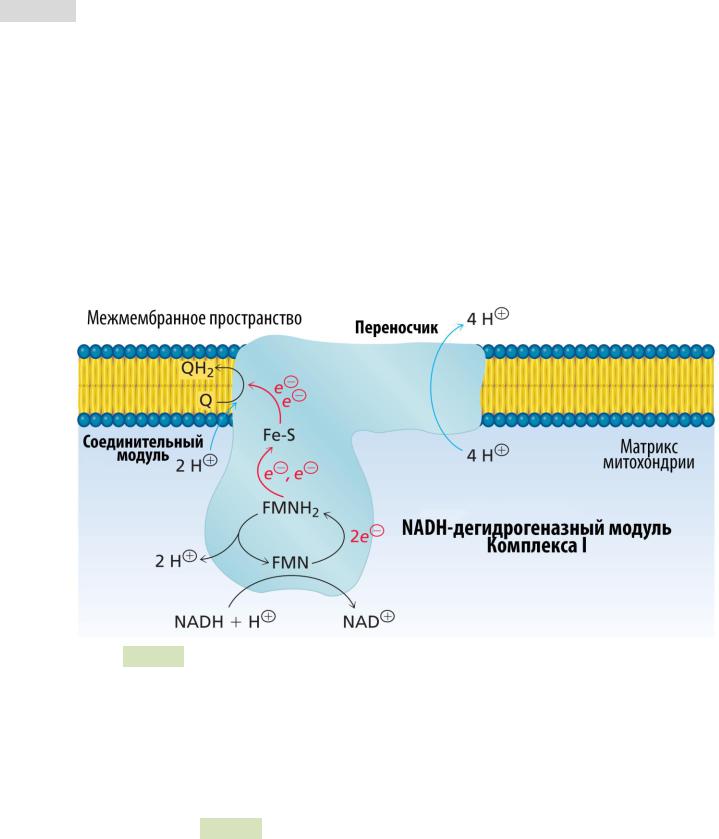
1.Комплекс I (NADH-CoQ 1 оксидоредуктаза; ~900 кДа, 45 уникальных субъединиц) включает (см. Рис. 120 ▼):
а. FMN (флавинмононуклеотид);
б. Ряд Fe-S центров (железистосерные центры).
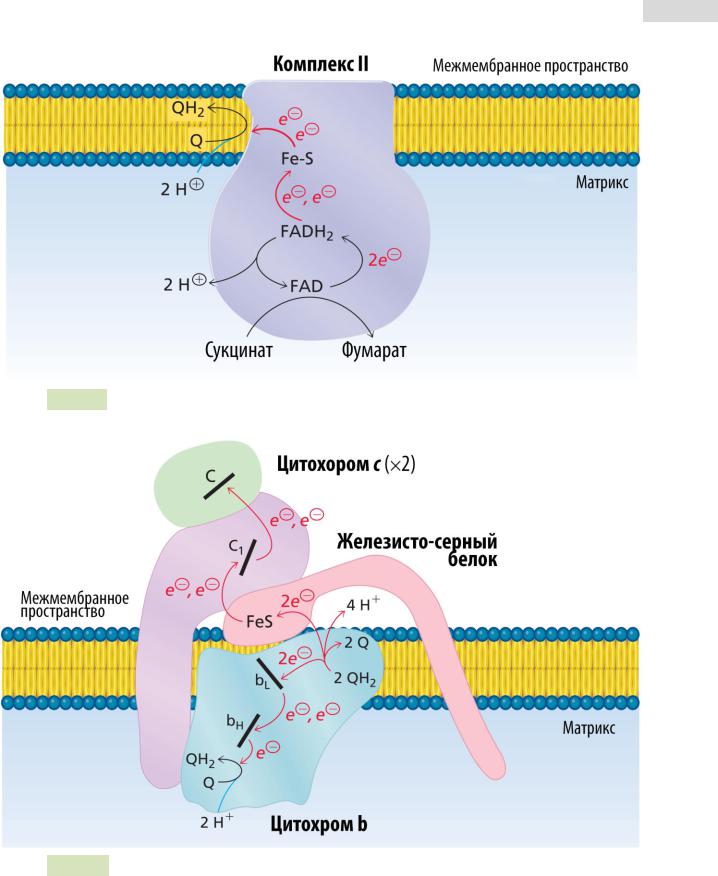
2.Комплекс II (сукцинат-CoQ оксидоредуктаза; ~420 кДа, 4 уникальные субъединицы) включает (см. Рис. 121 ▼):
а. FAD (флавинадениндинуклеотид); б. Ряд Fe-S центров;
в. Гем b560.
3.Кофермент Q (CoQ; единственный небелковый компонент дыхательной цепи).
4.Комплекс III (CoQ-цитохром с оксидоредуктаза; ~450 кДа, 9-11 уникальных субъединиц) включает (см. Рис. 122 ▼):
а. Гем bH (b562)
1 CoQ (англ. coenzyme Q – кофермент Q или просто Q) — убихинон. Может находиться в окисленной (Q) и восстановленной форме (QH2).
Сопряжение дыхания и окислительного фосфорилирования 203
б. Гем bL (b566) в. Fe-S центр; г. Гем c1.
5.Цитохром c («с» чит. как «цэ»).
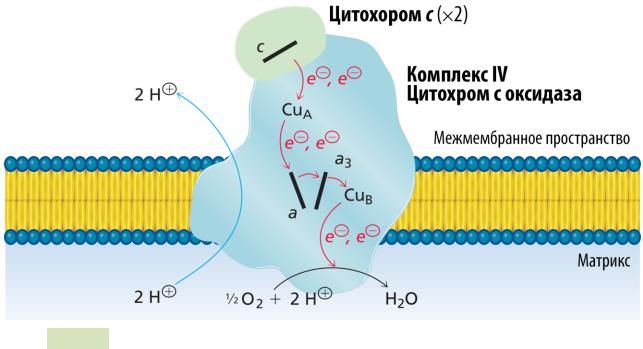
6.Комплекс IV (цитохром с оксидаза; димер ~410 кДа, 8-13 уникальных субъединиц) включает
(см. Рис. 123 ▼):
а. Гем a;
б. CuA и CuB (медные центры). в. Гем a3.
Сдыхательной цепью связан (но не от-
носится к ней) комплекс V (АТФ-синтаза), непосредственно синтезирующий АТФ.
Переносчиками электронов на дыхательную цепь выступают NADH (никотина- мид-аденин-динуклеотид) и FADH2 (флавин- аденин-динуклеотид). Эти соединения восстанавливаются в ходе разных метаболических путей и циклов (например, в гликолизе, окислительном декарбоксилировании пирувата, ЦТК). Схема переноса электронов изображена на рисунке.
11.3 Сопряжение дыхания и окислительного фосфорилирования
Питер Митчелл предложил хемиосмотическую теорию в 1961 году. Лишь спустя 10 лет его теория получила должное внимание и оценку. В 1978 году ему была вручена Нобелевская премия по химии.
Согласно теории Митчелла, свободная энергия переноса электронов запасается путём переноса протонов H+ из матрикса митохондрий в межмембранное пространство (и тем самым «запасается») и образования таким образом электрохимического градиента протонов H+. Электрохимический протенциал этого градиента протонов используется для синтеза АТФ. В этом и заключается основной смысл со-
пряжения дыхания и окислительного фосфори-
лирования. Некоторые ключевые наблюдения, объясняемые этой теорией:
Рис. 119. Дыхательная цепь митохондрий.
204Глава 11 Энергетический обмен
1.Окислительное фосфорилирование требует интактной митохондриальной мембраны.
2.Внутренняя митохондриальная мембрана непроницаема для ионов (H+, OH–, K+, Cl–), чья диффузия бы нарушила электрохимический градиент.
3.Транспорт электронов сопряжён с переносом протонов из матрикса в межмембранное пространство.
4.Соединения, повышающие проницаемость внутренней митохондриальной мембраны для протонов и тем самым нарушающие электрохимический градиент, не препятствуют транспорту электронов, однако ингибируют синтез АТФ; то есть они «разобщают» процессы переноса электронов и окислительного фосфорилирования.
Рис. 120. Перенос электронов и протонов Комплексом I
Комплексы I, III и IV дыхательной цепи генерируют протонный градиент, транспортируя протоны H+ из матрикса в межмембранное пространство одновременно с переносом электронов. Этот градиент ещё называют протондвижущей си-
лой.
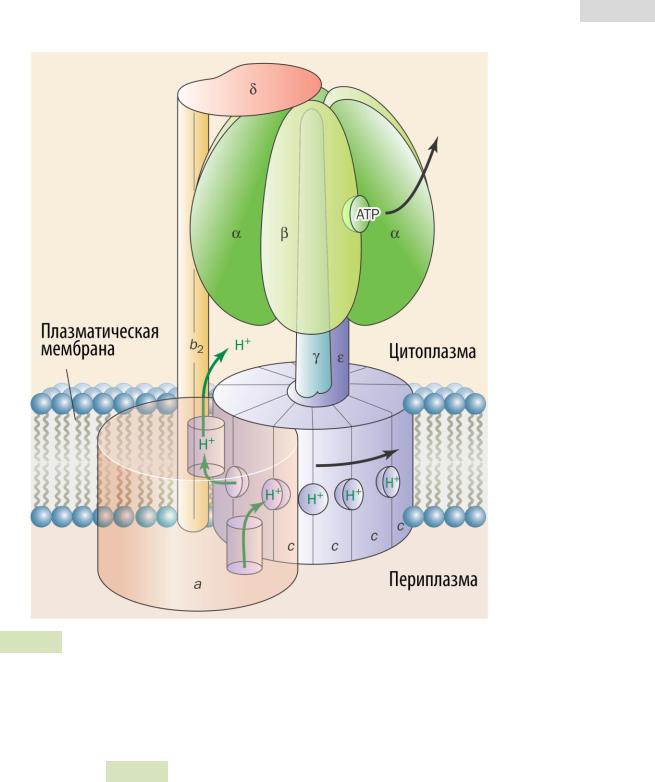
АТФ-синтаза (другие названия: F1F0-синтаза, Комплекс V) представляет собой полисубъединичный трансмембранный белок (фермент) с общей молекулярной массой 450 кДа (см. Рис. 124 ▼). Этот белок состоит из 2 частей:
1.F0 встроен в мембрану и состоит из 8 субъединиц.
2.F1 является периферическим мембранным белком из 5 типов субъеди-
ниц: α3β 3γ δ ε .
Сопряжение дыхания и окислительного фосфорилирования 205
Рис. 121. Перенос электронов Комплексом II
Рис. 122. Перенос электронов и протонов Комплексом III
206 |
Глава 11 |
Энергетический обмен |
Рис. 123. Перенос электронов и протонов Комплексом IV
Синтез АТФ можно представить в виде трёх фаз:
1.F0 осуществляет транслокацию протонов из межмембранного пространства в матрикс митохондрий.
2.F1 катализирует формирование фосфоангидридной связи (синтез АТФ из АДФ и Фн).
3.Сопряжение рассеяния протонного градиента с синтезом АТФ требует
взаимодействия F1 и F0.
Механизм синтеза — механизм разностного связывания — был предложен Полом Бойером (Paul Boyer). Согласно нему, у компонента F1 есть 3 каталитических протомера (состоящих из αβ -субъединиц), каждый из которых находится в особой конформации:
1.Слабо связывающая субстраты и продукты (L-состояние — англ. loose);
2.Прочно связывающая субстраты и продукты (T-состояние — англ. tight);
3.Свободная конформация, не связанная ни с какими молекулами (O-со- стояние — англ. open).
Сопряжение дыхания и окислительного фосфорилирования 207
Рис. 124. Модель АТФ-синтазы (Комплекса V) кишечной палочки E. coli
Свободная энергия переноса протонов используется для перехода между этими 3 состояниями. Фосфоангидридные связи в АТФ синтезируются только T- протомером, т.е. протомером, находящимся в T-состоянии. Отщепление синтезированной молекулы АТФ осуществляется от протомера в O-состоянии. Эта реакция происходит в 3 этапа (см. Рис. 125 ▼):
1.АДФ и Фн связываются с L-протомером (β DP);
2.Свободная энергия переноса протонов способствует изменению конформации L-сайта в T-сайт и синтезу АТФ в T-сайте (β TP). На этом этапе меняется конформация и других сайтов: T-сайт трансформируется в O- сайт (и происходит диссоциация молекулы АТФ), а O-сайт в L-сайт. Та-
ким образом, в любом момент времени существует 3 протомера (3 сайта, имеющих разную конформацию — T, L и O);
3.АТФ синтезируется в T-сайте, а диссоциирует в O-сайте (β E). Реакция синтеза АТФ находится в равновесии в активном центре фермента. Свободная энергия, обеспечиваемая потоком протонов, способствует от-