


Регуляция экспрессии генов у эукариот 139
«позволяет» или «не позволяет» РНК-полимеразе связаться с нужным промотором. Если связывания с промотором не произошло, то белок с гена, контролируемого данным промотором, синтезироваться не будет. Распространена гипотеза сменных σ- субъединиц: в зависимости от того, к какому промотору следует присоединиться РНКполимеразе, в коровой части фермента будет присутствовать σ-субъединица то одного, то другого типа (так же, как мы меняем насадки миксера или пылесоса при выполнении разной работы). Нуклеотидные последовательности промоторов, опознаваемых разными σ-субъединицами, существенно различаются и, как правило, не могут быть распознаны более чем одной σ-субъединицей. Например, у бактерии Bacillus subtilis один тип σ-субъединиц распознает последовательность TTGACA, расположенную в положении –35 и последовательность TATAAT в положении –10. Для распознавания другим типом σ-субъединиц этой же бактерии указанные последовательности должны быть ATATT и ATACA соответственно.
Транскрипцию практически всех жизненно важных генов, обеспечивающих гомеостатические клеточные процессы (репликацию ДНК, транскрипцию, трансляцию и т.д.), обеспечивают σ-субъединицы, называемые основными. Остальные процессы требуют альтернативных σ-субъединиц, которых в настоящее время описано не менее десяти. Например, у E. coli при резком повышении температуры (тепловом шоке) начинает синтезироваться альтернативная субъединица σ32 (с молекулярной массой 32 кДа). Ее присутствие придает РНК-полимеразе способность находить промоторы генов, кодирующие защитные белки — белки теплового шока.
7.2Регуляция экспрессии генов у эукариот
АОбщая характеристика
Общий принцип регуляции экспрессии генов у прокариот применим и к эукариотам: экспрессия отдельных генов может быть ингибирована или активирована с помощью особых белков, которые способны связываться с ДНК или РНК. Как и у прокариот, большинство регуляторных механизмов действуют на уровне транскрипции. Однако эукариотические организмы имеют значительно более сложную регуляцию экспрессии генов, чем прокариотические. Основные отличия связаны в первую очередь со строением ДНК:
1.Эукариотическая ДНК имеет огромные размеры по сравнению с прокариотической. К примеру, кишечная палочка E. coli имеет одну кольцевую хромосому (4 миллиона нуклеотидных пар), а самый примитивный эукариотический организм — Saccharomyces cerevisiae — имеет 16 хромосом, и длина его ДНК — 16 миллионов нуклеотидных пар.
2.Генотип у эукариот в большинстве случаев диплоидный.
3.Эукариотическая ДНК связана с белками гистонами и образует хроматин. Хроматин состоит из повторяющихся элементов — нуклеосом (см. тему 3). Каждая молекула гистона в нуклеосоме имеет N- терминальный «хвост», который может ковалентно модифицироваться. Именно эта модификация гистонов играет значимую роль в регуляции экспрессии эукариотических генов.
140Глава 7 Регуляция биосинтеза белка
4.Эукариотические гены не организованы в опероны. Гены, кодирующие белки, участвующие в одном метаболическом пути, часто находятся в разных частях генома. Однако у эукариот существует кластеризация генома — некоторые гены объединены в кластеры.
5.Жизненный цикл у эукариот сложнее и продолжительнее, чем у прокариот.
6.Процессы транскрипции и трансляции у эукариот разделены в пространстве и во времени.
Некоторые особенности регуляции экспрессии генов у эукариот:
1.У эукариот большинство генов «выключено», т.е. транскрипция на них не идёт, поэтому регуляция очень часто направлена на их «включение». Постоянно транскрибируется не более 7-10% генов.
2.Регуляция осуществляется очень «гибко». Возможна различная интенсивность синтеза белка (низкая или высокая).
3.Транскрипционный аппарат состоит из множества белковых факторов, комбинирование которых делает регуляцию более разнообразной.
4.Эукариоты активно нарабатывают мРНК заранее и используют их «по потребности».
БСтруктура хроматина и регуляция экспрессии генов
Большая часть ДНК в многоклеточных организмах не экспрессируется: к ней относятся участки, не кодирующие белок или РНК, а также гены, чья экспрессия в клетках определённого типа нежелательна. Несмотря на то, что все клетки организма содержат одинаковый набор молекул ДНК, их экспрессия тканеспецифична. Например, многие клетки поджелудочной железы синтезируют и секретируют пищеварительные ферменты, при этом островковые клетки поджелудочной железы синтезируют инсулин или глюкагон.
Неэкспрессируемая ДНК — гетерохроматин — обычно высокоспирализована и потому недоступна для РНК-полимераз (см. Рис. 93 ). Транскрипционно активная форма ДНК — эухроматин — имеет меньшую степень спирализации.
Для того, чтобы экспрессировать генетическую информацию ДНК, транскрипционный аппарат клетки должен получить «доступ» к ДНК, которая упакована в нуклеосомную нить гистоновыми белками. Нуклеосомы не являются фиксированными структурами — они могут подвергаться перестройке. Например, гистоны способны ковалентно модифицироваться.
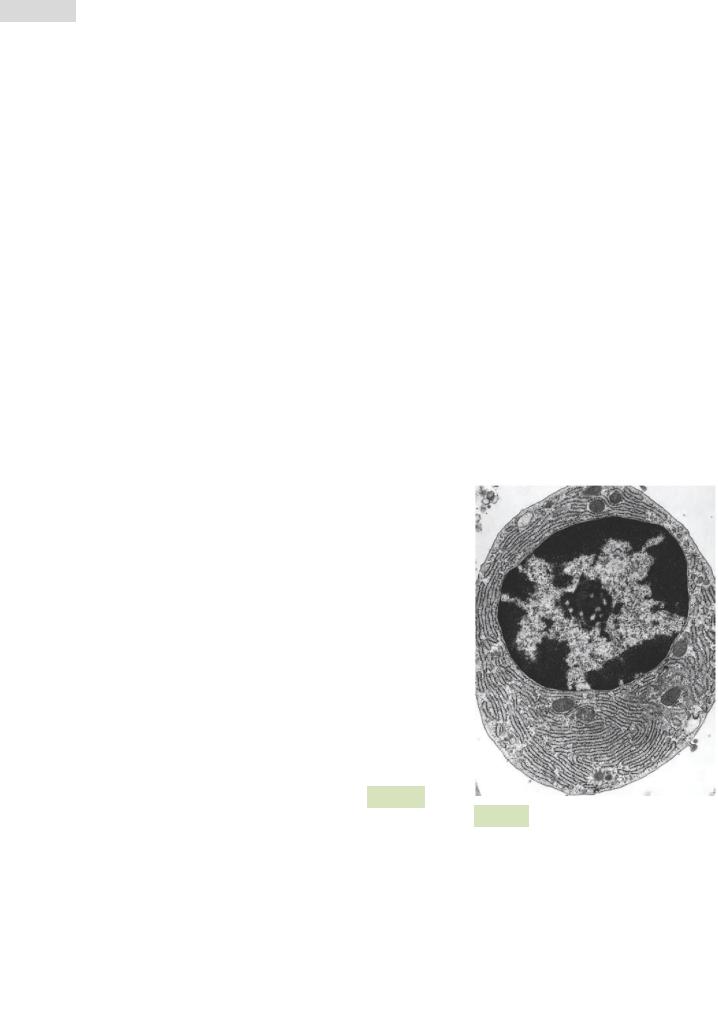
Рис. 93. Электронная микрофотография B-лимфоцита. Значи-
тельную часть клетки занимает ядро, где темные участки — гетерохроматин (неактивная ДНК), а светлые — эухроматин (активная ДНК). В центре ядра — ядрышко. Обратите внимание на обилие шероховатого эндоплазматического ретикулума в клетке.
Регуляция экспрессии генов у эукариот 141
Хроматин-перестраивающие комплексы
В ходе матричных процессов — транскрипции и репликации — полимеразные белки и белковые факторы должны иметь возможность присоединяться к цепи ДНК. Однако практически вся ДНК у эукариот связана с гистонами и образует нуклеосомную нить. В середине 90-х годов стало известно, что хроматин содержит комплексы,
перестраивающие нуклеосомы и использующие энергию АТФ. Эти комплексы разры-
вают связи между ДНК и гистонами, благодаря чему ДНК становится доступной для других белков, а гистоновый октамер «скользит» вдоль цепи ДНК или даже перено-
сится на другую цепь. Таким образом, хроматин-перестраивающие комплексы
придают хроматину «плавающую» структуру, не меняя общую степень упаковки ДНК. Хроматин-перестраивающие комплексы состоят из множества субъединиц, организованных в комплекс SWI/SNF (~11 субъединиц) и комплекс RSC. Донором энергии служат молекулы АТФ. Предполагаемый механизм действия этих комплексов заключается во внесении локальных изгибов в цепь ДНК, благодаря созданию «волн», которые постепенно разрывают связи между ДНК и гистонами. В результате нуклеосомный кор «соскальзывает» с нужного участка ДНК, приоткрывая его для бел-
ков-ферментов.
Архитектурные белки высокомобильной группы
Среди негистоновых белков, связанных с ДНК, самыми распространенными являются белки высокомобильной группы (HMG-белки). Они были названы так потому, что обладают высокой электрофоретической мобильностью в полиакриламидном геле (электрофорез). Эти белки имеют малую массу ~30 кДа и необычный аминокислотный состав: ~25% осно́вных и 30% кислых аминокислот.
HMG-белки способны изменять конформацию ДНК, вызывая скручивание, выпрямление, разматывание цепи или формирование петель.
Ковалентная модификация гистонов
Гистоны способны посттрансляционно модифицироваться:
—Ацетилирование по определённым остаткам лизина (присоединение ацетильной группы —CH3CO).
—Метилирование определённых остатков лизина и аргинина (присоединение метильной группы —CH3).
—Фосфорилирование определенных остатков серина и треонина (присоединение остатка фосфорной кислоты —PO4 3–).
—Убиквитинилирование определённых остатков лизина.
Все модификации за исключением метилирования снижают заряд гистонов (он становится отрицательным), поэтому их взаимодействие с ДНК нарушается (ДНК заряжена отрицательно, а гистоны в норме — положительно). Гистоны отщепляются от ДНК, и транскрипция начинается.
Метилирование гистонов H3 и H4 по остаткам лизина и аргинина выключает экспрессию генов, вызывая формирование гетерохроматина.