

Врожденный (неспецифический) иммунитет 163
Результат лектинового пути: формируется активный комплекс C4b2a; C3b связывается с мембраной патогена.
Е Классический путь активации комплемента
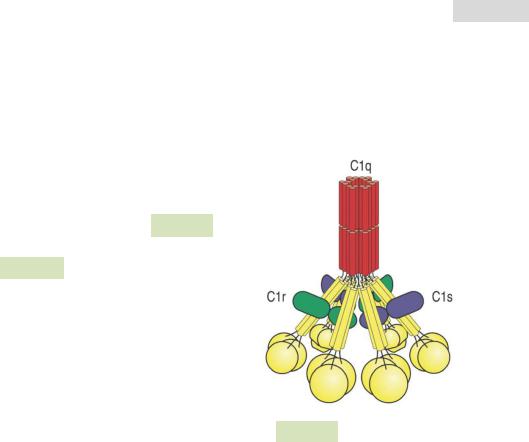
Классический путь активации комплемента напоминает лектиновый: он тоже использует «молекулусенсор» для узнавания патогенов, но в этом пути эту роль играет комплекс C1 или просто C1 (см. Рис. 100 ). Этот комплекс взаимодействует как с самим патогеном, так и с антителами (см. Рис. 101 ▼), поэтому классический путь активации комплемента участвует не только в неспецифическом иммунном ответе, но и в специфиче-
ском. Комплекс C1 состоит из большого белка C1q, узнающего патогены, и двух неактивных сериновых протеаз
— C1r и C1s:
Этапы активации:
1.Комплекс C1 связывается с лигандом (патогеном или комплексом антитело + патоген). Активируется C1r, который переводит про-
теазу |
C1s |
в |
активное |
состояние: |
C1 + лиганд → C1r C1s*. |
|
|||
Рис. 100. Строение комплекса C1.
2.C1s расщепляет C4 до C4a и C4b. Последний связывается с мембраной и
C2.
3.C1s расщепляет C2 до C2a и C2b. Образуется активный комплекс C4b2a
(C4b + C2a).
4.Комплекс C4b2a — это активная C3-конвертаза, расщепляющая C3 на C3a и C3b. C3b связывается с мембраной патогена.
Результат классического пути: формируется активный комплекс C4b2a; C3b связывается с мембраной патогена.
164 |
Глава 9 |
Иммунитет и антитела |
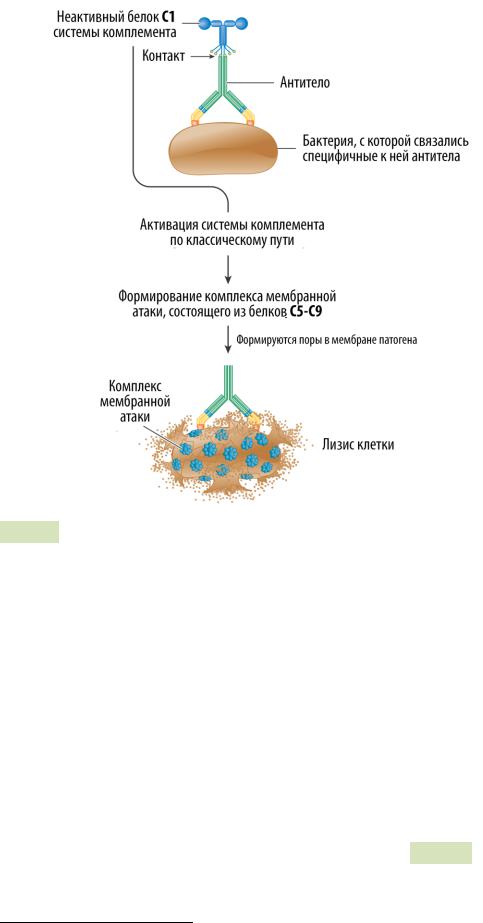
Рис. 101. Активация системы комплемента по классическому пути комплексом антиген-антитело.
Ж Альтернативный путь активации комплемента
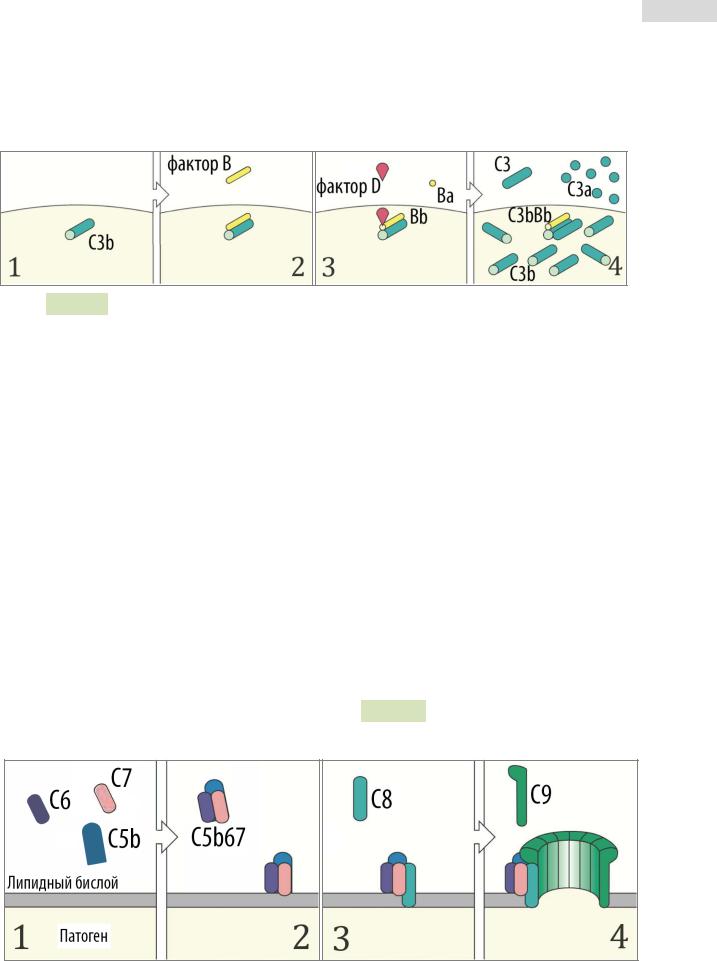
Этапы активации:
1.Белок C3 («си-три» или «це-три») попадает в кровь из печени и время от времени спонтанно активируется (C3*) с помощью гидролиза тиоэфирной связи внутри молекулы.
2.Активированный C3* связывается с фактором B.
3.Фактор D расщепляет фактор B на факторы Ba и Bb.
4.Комплекс C3*Bb представляет собой нестабильную C3-конвертазу13 , расщепляющую неактивные белки C3 на C3a и C3b. Последний быстро инактивируется, если не контактирует с мембранами патогенов. Однако, если такой контакт происходит, C3b связывается с фактором B, который снова расщепляется фактором D на Ba и Bb (см. Рис. 102 ▼).
5.Образуется комплекс C3bBb — он является уже стабильной C3конвертазой, расщепляющей неактивные белки C3 на C3a и C3b. С3b
13C3-конвертаза стабильна лишь некоторое время, однако она стабилизируется белком пропердином (фактор P), который секретируют нейтрофилы в присутствии патогенов.
Врожденный (неспецифический) иммунитет 165
быстро покрывают поверхность мембраны патогена. Происходит активация терминальных компонентов комплемента (см. раздел 10).
Результат альтернативного пути: формируется активный комплекс C3bBb;
C3b связывается с мембраной патогена.
Рис. 102. Активация комплемента по альтернативному пути.
З Активация терминальных компонентов комплемента
Все три пути активации комплемента сходятся на этапе образования C3конвертазы. Следующим этапом является формирование активной C5-конвертазы.
1. C5-конвертаза формируется так:
а. В классическом и лектиновом пути C4b2a связывается с C3b. Образуется C4b2a3b.
б. В альтернативном пути C3bBb связывается с C3b. Образуется
C3b2Bb.
Комплексы C4b2a3b и C3b2Bb представляют собой активные C5-
конвертазы.
2.C5-конвертаза связывается с белками C5 и расщепляет их на C5a и C5b.
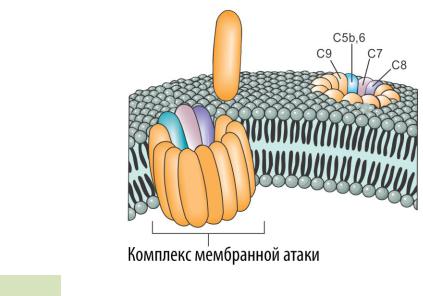
3.C5b связывается с C6.
4.Комплекс C5b6 (C5b + C6) связывается с C7.
5.Далее происходит последовательное связывание комплекса C5b67 с белками C8 и C9.
6.Ключевую роль в формировании комплекса мембранной атаки играют 10-16 молекул белка C9 (см. Рис. 103 ▼). Именно они и образуют мембранную литическую пору.
166 |
Глава 9 |
Иммунитет и антитела |
Рис. 103. Формирование комплекса мембранной атаки.
И Как фагоциты отличают чужеродные клетки от «своих»?
Безусловно, фагоциты должны отличать патогенные клетки от клеток своего организма. Для этого существует два механизма:
1.У фагоцитов на поверхности мембран находятся особые Толл-подоб- ные рецепторы. Эти рецепторы «узнают» вещества, содержащиеся в мембранах патогенов — молекулярные особенности патогена, называе-
мые PAMP (англ. pathogen-associated molecular patterns). Наиболее из-
вестными из них являются липополисахариды (ЛПС) грамотрицательных бактерий и пептидогликаны клеточных стенок грамположительных бактерий. Взаимодействие Толл-подобных рецепторов с патогенами (или чужеродными веществами) стимулирует секрецию фагоцитирующими клетками цитокинов (регуляторных молекул):
а. Привлекают к месту обнаружения патогена другие клетки имму-
нитета — нейтрофилы и моноциты, а также T- и B-лимфоциты;
б. Активируют другие механизмы врожденного иммунитета — фа-
гоцитоз и лихорадку.
2.В организме человека синтезируются особые вещества, «помечающие» патогены для фагоцитирования — опсонины. Важнейшие опсонины —
антитела и белки системы комплемента (например, белки C3b). Опсо-
нины обеспечивают более тесный и надёжный контакт между фагоцитом и патогеном.
КВещества, секретируемые фагоцитами
Наиболее важные вещества, секретируемые фагоцитами:
1.Макрофаги секретируют оксид азота NO, токсичный для бактериальных клеток.