

Эксцизионная репарация оснований (BER) 151
8.6 Эксцизионная репарация оснований (BER)
BER-система исправляет следующие ошибки в ДНК: 8-оксогуанин, урацил (образуется при дезаминировании цитозина или его случайной вставке в ДНК вместо тимина).
Стадии репарации (см. Рис. 94 ):
1.Поврежденное азотистое основание обнаруживается и удаляется ферментом ДНК-гликозилазой. В
результате образуется АП-сайт (апуриновый или апиримидиновый), т.е. нуклеотидный остаток без азотистого основания (сахарофосфатная группа).
2.В цепь ДНК вносится разрыв вблизи АП-сайта ферментом АП-
эндонуклеазой.
3.Сахарофосфатная группа — дезоксирибоза и остаток фосфорной кислоты — и несколько следующих нуклеотидных остатков вырезаются ферментом экзонуклеазой.
4.Брешь застраивается ДНК-поли-
меразой I и сшивается ДНК-лига- зой.
8.7Эксцизионная репарация нуклеотидов (NER)
NER-система у человека достаточно эффективно исправляет последствия действия на организм канцерогенов, солнечных лучей и табачного дыма.
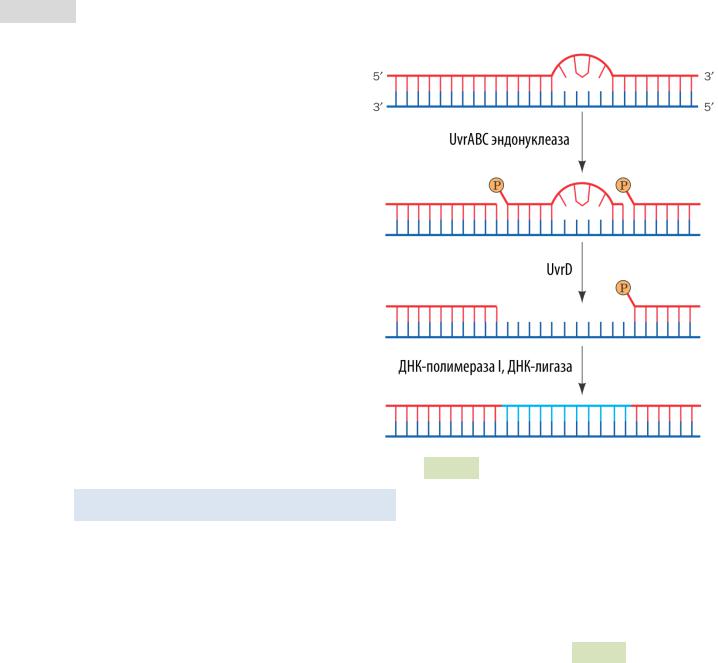
Эксцизионная репарация нуклеотидов у кишечной палочки E. coli — АТФ-зависимый процесс, в котором участвуют ферменты UvrA, UvrB, UvrC. Эту систему ещё часто называют
UvrABC-эндонуклеазой (см. Рис. 95 ▼).
Рис. 94. Механизм BER-репарации:
в данном случае удаляется лишь один нуклеотид
152Глава 8 Мутации и репарация
Стадии репарации:
1.Ферментный комплекс UvrABC разрезает цепь ДНК у 7-го и 3-го/4-го нуклеотида с 5’- и 3’- стороны от повреждения, соответственно.
2.Вырезанный 11-12 нуклеотидный фрагмент затем вытесняется фер-
ментом UvrD (хеликаза
II).
3.ДНК-полимераза I син-
тезует комплементарную цепочку, а ДНК-ли- газа сшивает цепи.
Уэукариот NER-система состоит из 16 белков и вырезает отрезок длиной примерно 30 нуклеоти-
дов. |
Рис. 95. Механизм NER-репарации |
|
8.8Мисметч репарация
Входе репликации ДНК-полимераза может совершать ошибки. Неспаренные нуклеотиды удаляются с помощью мисметч репарации (англ. mismatch — несовпадение). MMR-система способна исправить инсерции и делеции до 4 нуклеотидов. Важность MMR-репарации подчеркивается тем фактом, что у людей с наследственными дефектами этой системы наблюдается повышенная предрасположенность к раковым заболеваниям. Стадии репарации у кишечной палочки E. coli (см. Рис. 96 ▼):
1.Белок MutS связывается с некомплементарной парой нуклеотидов.
2.К MutS присоединяется белок MutL.
3.Комплекс MutS2-MutL2 транслорцируется вдоль цепи ДНК таким образом, что формируется петля, замыкающаяся на комплекс MutS2-MutL2.
4.При движении по цепи ДНК комплекс MutS2-MutL2 распознает неметилированную цепочку нуклеотидов GATC и связывается с белком MutH
(эндонуклеаза).
5.MutH активируется и разрезает цепь с 5’-конца у неметилированной цепочки GATC (так система распознает новосинтезированную дочернюю цепь ДНК — после синтеза она лишь частично метилирована, а материнская цепь метилирована полностью). Такая неметилированная последовательность GATC может находиться и в 1000 нуклеотидных остатках от сайта повреждения.
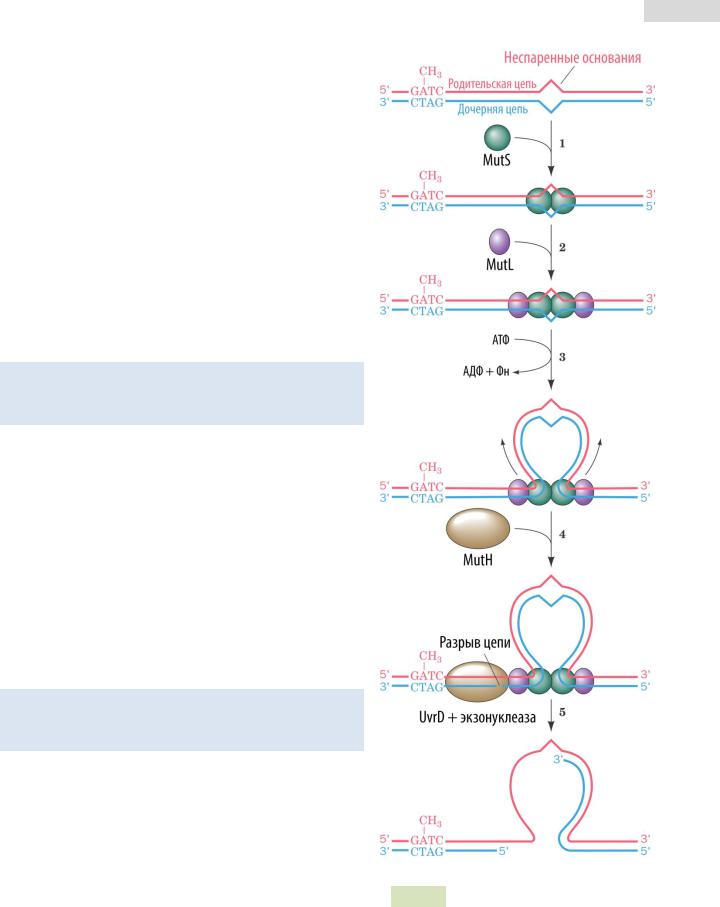
Репарация двунитевых разрывов 153
6.Хеликаза UvrD (из NER-системы) разрывает водородные связи между родительской и дочерней цепями.
7.Экзонуклеаза полностью расщепляет дефектный участок ДНК.
8.ДНК-полимераза III синтезирует участок дочерней цепи, комплементарный родительской ДНК.
Учеловека нет гомологов белка MutH, поэтому используется другой механизм распознавания дочерней цепи. По одной из теорий, дочерняя цепь определяется по разрывам между фрагментами Оказаки.
8.9Репарация двунитевых разры-
вов
Двунитевые разрывы в ДНК появляются из-за воздействия ионизирующей радиации или свободных радикалов (продуктов окислительного метаболизма). Нерепарированные двунитевые разрывы могут стать летальными для клетки или спровоцировать рак. Существует два основных механизма репарации двунитевых разрывов:
1. Рекомбинационная репарация;
2. Негомологичное соединение концов.
8.10 Негомологичное соединение цепей ДНК при двунитевых разрывах
Негомологичное соединение концов ДНК |
|
|
при двунитевых разрывах требует отщепления |
|
|
нескольких нуклеотидов, поэтому часто тоже |
|
|
провоцирует мутации, но они не так опасны |
|
|
для клетки, как нерепарированные двунитевые |
|
|
разрывы. В этом виде репарации участвуют |
Рис. 96. Механизм MMR-репарации. |
|
белки Ku (димер Ku70 + Ku80). Они присоеди- |
||
|
||
няются к ДНК в области малой и большой бо- |
|
роздок и отщепляют несколько нуклеотидных остатков. Затем происходит сшивание цепей. К 70-и годам в соматических клетках человека скапливается до 2000 подобных «шрамов» от негомологичного соединения цепей ДНК.
154 |
Глава 8 |
Мутации и репарация |
8.11 SOS-репарация (SOS-ответ)
Агенты, повреждающие ДНК, индуцируют комплексную систему защиты в клетках кишечной палочки E. coli — SOS-ответ. Эта система регулируется белками LexA (репрессор) и RecA (ДНК-связывающий белок). В норме LexA подавляет экспрессию гена SOS. Однако при повреждении цепи ДНК связываются с RecA и формируют комплекс, который выключает LexA. SOS-система контролируется 43 генами, среди которых — гены, кодирующие ДНК-полимеразы IV и V. Эти полимеразы способны синтезировать цепь, не имея матрицы в виде комплементарного участка ДНК. Поэтому SOS-репарация совершает ошибки и является мутагенной, однако позволяет продлить срок жизни клетки. По мнению некоторых учёных, эта система позволяет клеткам приобрести новые признаки, утратив при этом какие-либо другие.
8.12 Рекомбинационная репарация
Рекомбинационная репарация у кишечной палочки осуществляется следующим образом:
1.Репликативная вилка встречает одноцепочечный разрыв и останавливается.
2.Процесс восстановления начинается с переноса белками RecBCD + RecA участка новосинтезированной и неповрежденной ДНК на двуцепочечную ДНК в место повреждения (с 3’-конца).
3.Образуется структура Холлидея. Происходит обмен 3’-концами между репликативными вилками с помощью белков RuvAB.
4.Белок RuvC разрывает структуру Холлидея и восстанавливает репликативные вилки.
5.Процесс репликации продолжается при участии особой праймосомы.
Термины
Мутация, ДНК гликозилазы, АП-сайт, АП-эндонуклеазы, ДНК фотолиазы, SOSответ, рекомбинационное восстановление ДНК, транспозоны.
Вопросы к занятию
1.Мутации и мутагены. Виды мутаций и механизмы их возникновения.
2.Канцерогенез.
3.Репарация: классификация типов репарации, прямая и непрямая репарация.
4.Эксцизионная репарация:
a.Репарация нуклеотидов (NER);
b.Репарация оснований (BER);
c.Мисметч-репарация (MMR).
5.Рекомбинация как одна из форм репарации.
6.SOS-репарация.