

16Глава 1 Аминокислоты и белки
—Аминокислоты, образующие β -листы: серин, гистидин, цистеин, треонин,
тирозин, фенилаланин, триптофан, изолейцин, валин.
—Аминокислоты, образующие повороты (шпильки): глицин, аспарагин, аспар-
тат, пролин, серин.
В Третичная структура белка
Третичная структура белка — это общее пространственное расположение всех атомов в молекуле белка (см. Рис. 12 ▼). Химические связи, стабилизирующие третич-
ную структуру: координационные связи (с ионами металлов), гидрофобные взаимодей-
ствия (между неполярными аминокислотами), дисульфидные связи (–S–S–), ионные связи (между заряженными радикалами) и водородные связи (участвуют как радикалы, так и пептидные группы).
Таблица 2 Химические связи и их энергии
|
Тип химической связи |
|
|
Примеры |
|
|
|
Энергия связи |
|
|
|
|
|
|
|
(кДж/моль) |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O — H |
|
|
|
460 |
|
Ковалентные |
|
|
C — H |
|
|
|
414 |
|
|
|
|
|
|
C — C |
|
|
|
348 |
|
Нековалентные |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|||
1. |
Ионные связи |
|
|
|
|
86 |
|
||
|
|
|
|
|
|
|
|
|
|
2. |
Силы ван-дер-Ваальса |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
— Водородные связи |
|
|
|
|
20 |
|
||
|
|
|
|
|
|
|
|
|
|
|
— Диполь-дипольные |
|
|
|
|
9,3 |
|
||
|
взаимодействия1 |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
||
|
— Лондоновские |
|
|
|
|
0,3 |
|
||
|
дисперсионные силы |
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
1 См. Приложение 1. Аминокислоты и белки
Уровни структурной организации белков 17
Третичная структура глобулярных белков представляет собой глобулу (или клубок). Большинство глобулярных белков растворимы в клетке. Как правило, ядро глобулярного белка содержит много неполярных аминокислотных остатков (валин, лейцин, изолейцин, фенилаланин, метионин), которые изолированы от водной среды и образуют гидрофобное ядро (гидрофобный эффект). Это ядро плотно упаковано, внутри него действуют слабые силы ван-дер-Ваальса
(обуславливающие гидрофобные взаимодействия). Могут воз-
никать водородные связи между полярными незаряженными радикалами (серин, треонин, аспарагин, глутамин, тирозин и триптофан). Заряженные полярные аминокислотные радикалы часто расположены на поверхности белка и формируют ионные пары (ионные связи) между собой (таким образом сближаются удаленные участки полипептидной цепи) или вступают в контакт с растворителем — молекулами воды (см. Рис. 13 ▼). Кроме того, заряженные радикалы часто связываются с неорганическими ионами (в т.ч. ионами металлов с образованием координационных связей), что снижает силы отталкивания между фрагментами цепи. Внутри полипептидной цепи и между цепями могут образовываться дисульфидные мостики (в их образовании участвуют сульфгидрильные группы цистеина, –S—S–).
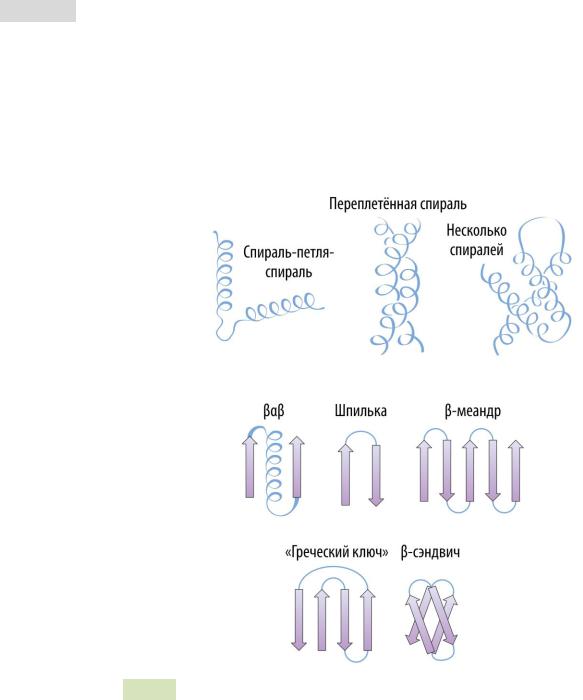
В рамках третичной структуры можно выделить неко-
торые надвторичные структуры: спираль-поворот-спираль,
переплетённая спираль, β αβ , шпилька, β -меандр и др. (см. Рис. 14 и Рис. 15 ▼).
Многие белки состоят из отдельных фрагментов, имеющих свою индивидуальную укладку, функцию и стабильность — доме́нов. Длина аминокислотной цепочки одного домена может составлять от 25 до 300 и более аминокислотных остатков.
Домены белков по своему составу и структуре классифицируют на 4 группы:
—«α»-белки содержат только α-спирали.
—«β»-белки — только β -структуры.
—«α/β»-белки состоят из параллельных β -структур
ипараллельных α -спиралей, которые антипараллельны друг другу.
—«α+β»-белки подразделяют на две класса:
Белки первого класса напоминают «α/β »- белки тем, что в них слой α-спиралей лежит на β -листе. Их α и β участки в цепи и в пространстве регулярно чередуются.
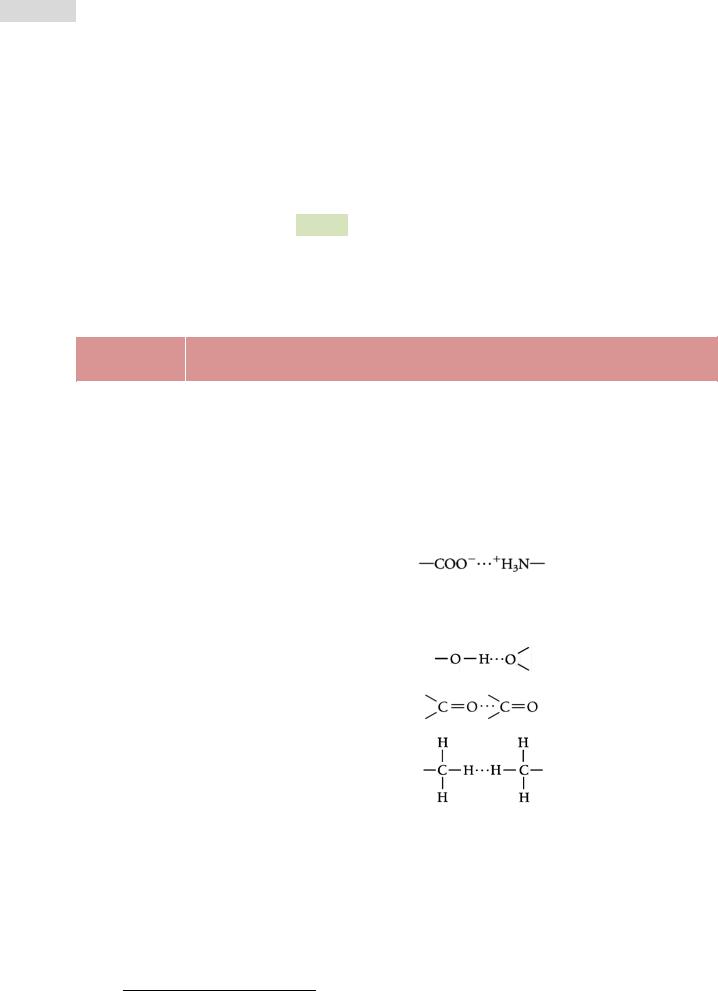
Рис. 12. Третичная структура цитохрома b562 кишечной па-
лочки E. coli (ленточ-
ная модель).
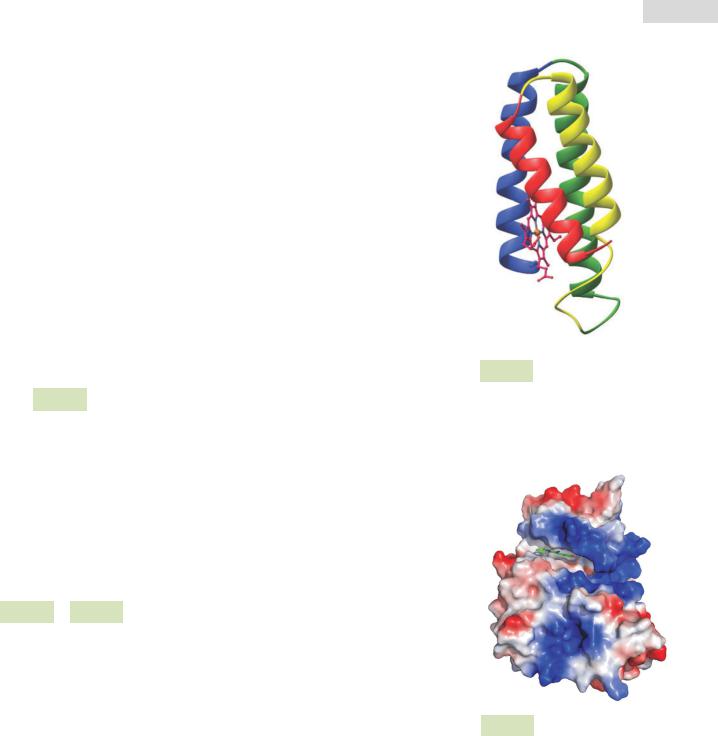
Рис. 13. Компьютерная визуализация молекулы протеинкиназы CK2 (белок-фермент): синим цветом обозначены положительно заряженные радикалы, красным цветом — отрицательно заряженные, а серым — нейтрально заряженные.
18Глава 1 Аминокислоты и белки
Белки другого класса («собственно» «α+β »-белки) не имеют такого чередования, в их цепях α-спирали пространственно удалены от β -
структур.
Стоит упомянуть, что некоторые белки вовсе лишены вторичной и упорядоченной третичной структур (или содержат неупорядоченные фрагменты). Это вызвано большим количеством заряженных аминокислот и пролина в их цепях.
Рис. 14. Надвторичные структуры белков.
ГАнализ и предсказание третичной структуры белка
Третичную структуру можно определить с помощью рентгеновской кристаллогра́фии, пропуская рентгеновские лучи через чистые белковые кристаллы, выращенные в определенных условиях, и затем анализируя дифракционную картину. Данная картина создаётся в результате рассеивания лучей электронами каждого атома молекулы. Рассеянные рентгеновские лучи могут взаимно усиливаться или ослабляться, создавая характерную картину для каждой молекулы.
Третичную структуру белка можно отчасти «предсказать», зная первичную структуру белка. Разработаны компьютерные алгоритмы, позволяющие сделать это, используя банк данных с уже известными конформациями белков. Другие методы основаны на анализе конформации отдельных участков молекулы: существуют данные о том, какие аминокислоты чаще всего образуют α-спирали и β -листы, а какие
Уровни структурной организации белков 19
наоборот препятствуют этому и не формируют регулярных структур. Эту информацию успешно используют при прогнозировании третичной структуры белка.
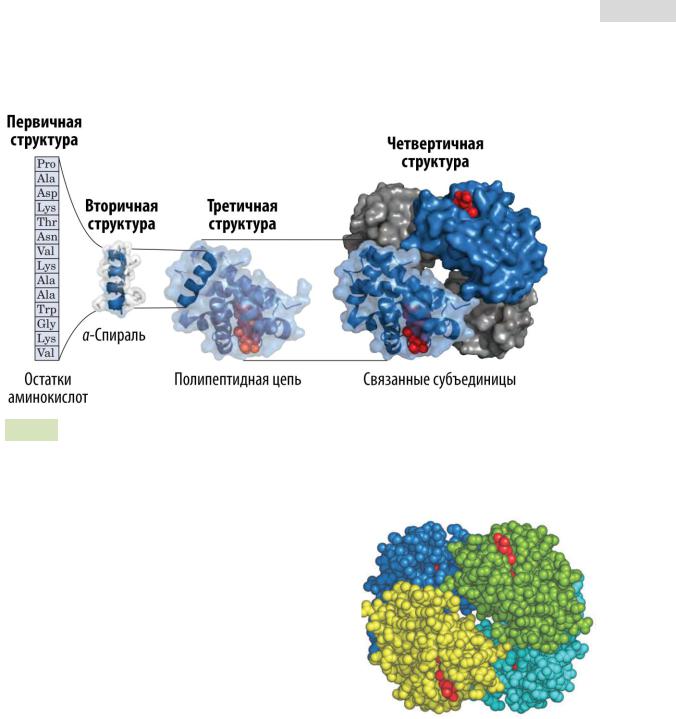
Рис. 15. Уровни структурной организации белков.
Д Четвертичная структура белка
Четвертичной структурой обладают лишь те белки, которые содержат две и более полипептидные цепи. Каждую цепь при этом называют субъединицей (или протоме́ром). Белок, состоящий из нескольких протомеров, называют олигомерным. Количество цепей может варьироваться от 2 до 10 и более. Чаще всего встречаются димерные (две цепи), тримерные (три цепи) и тетрамерные (четыре цепи) белки. Четвертичной структу-
рой обладает, например, белок гемоглобин (см. |
|
|
|
|
|
|
|
|
|
Рис. 16 |
). Он имеет 4 субъединицы (т.е. 4 |
|
|
|
|
Рис. 16. |
Четвертичная струк- |
||
полипептидные цепи). |
|
|||
|
тура гемоглобина. |
|||
Субъединицы в полисубъединичных бел- |
|
|||
|
|
|
||
ках связаны с помощью нековалентных связей: |
|
|
|
|
|
|
|
||
гидрофобных и электростатических взаимодействий. Поскольку эти связи достаточно слабые, субъединицы часто можно разделить в лабораторных условиях. Однако in vivo они остаются прочно связанными.
Некоторые факторы, способствующие образованию четвертичной структуры:
1.Олигомеры обычно более стабильны, чем их диссоциировавшие субъединицы. По всей вероятности, четвертичная структура продлевает «время жизни» белка.
2.Активные центры некоторых олигомерных ферментов формируются из аминокислотных остатков, относящихся к разным полипептидным цепям.