

120 |
Глава 6 |
Трансляция |
релиз-факторы (RF-белки). Они способствуют диссоциации рибосом на две субъединицы — малую и большую.
Д Этап 5. Фолдинг и посттрансляционная модификация
Любой полипептид должен принять нативную конформацию для достижения биологической активности. До или после фолдинга белок должен подвергнуться ферментативному процессингу — отщеплению нескольких аминокислот с конца цепи (обычно с N-конца), присоединению ацетильной, метильной, карбоксильной или другой химической группы (и часто не одной), протеолизу (отщеплению целой части цепи) или присоединению олигосахаридов или каких-либо других простетических групп.
6.4 Рибосомы
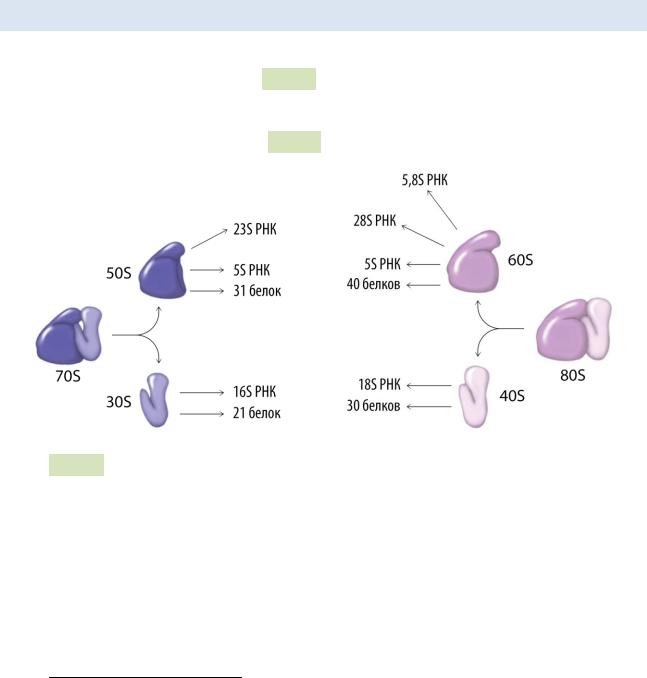
Рибосомы — это крупные рибонуклеопротеидные комплексы, состоящие из большой и малой субъединицы (см. Рис. 79 ▼). В состав рибосом входят рРНК и белки. Собранные и активные рибосомы имеют 3 сайта: А-сайт (сайт связывания с амино-
ацил-тРНК), Р-сайт (сайт, в котором находится пептидил-тРНК8) и Е-сайт (сайт, в
котором деацилируется тРНК) (см. Рис. 80 ▼).
Рис. 79. Строение про- и эукариотической рибосомы: слева — прокариотическая
70S рибосома (состоит из большой 50S и малой 30S субъединиц), справа — эукариотическая 80S рибосома (состоит из большой 60S и малой 30S субъединиц).
8 Пептидил-тРНК — транспортная РНК, связанная с синтезируемой пептидной цепью.
Инициация у прокариот 121
Рис. 80. Сайты связывания тРНК у прокариотической рибосомы.
6.5 Инициация у прокариот
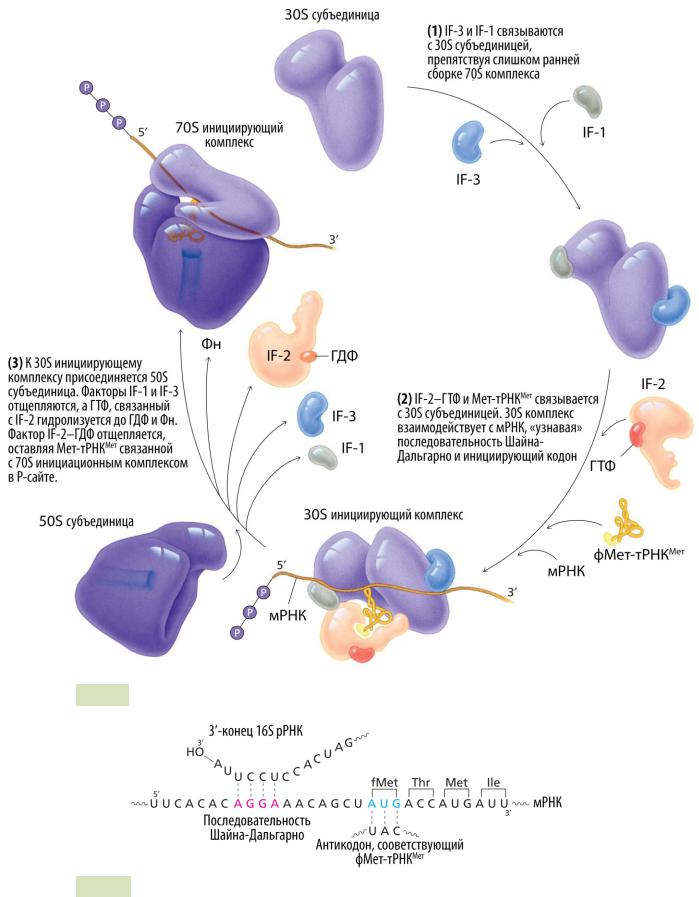
Инициация — это первая стадия трансляции, в ходе которой происходит сборка 70S рибосомы (инициаторного комплекса), узнавание старт-кодона AUG9 (кодирует аминокислоту формилметионин10), установка рамки считывания и связывание с формилметионил-тРНКМет (транспортная РНК, несущая остаток формилметионина)
вP-сайте рибосом. Инициация происходит в несколько этапов (см. Рис. 81 ▼):
1.Рибосомы находятся в диссоциированном состоянии перед трансляцией, то есть обе субъединицы разделены, и с 30S субъединицей рибосом связан фактор IF-3, препятствующий сборке 70S комплекса.
2.Факторы IF-1, IF-2–ГТФ и мРНК связываются с 30S субъединицей в произвольном порядке. Связывание формилметионил-тРНК с 30S субъединицей происходит не по принципу кодон-антикодоновой комплементарности, а благодаря взаимодействию с фактором IF-2. Фактор IF-3 способствует связыванию мРНК с 30S комплексом. Затем 16S рРНК малой субъединицы ри-
босом «находит» на мРНК последовательность Шайна-Дальгарно (5’ –
GGAGGU– 3’), связывается с ней по принципу комплементарности и устанавливает рамку считывания на инициирующий кодон AUG (см. Рис. 82 ▼).
3.50S субъединица присоединяется к 30S инициаторному комплексу. Факторы IF-1 и IF-3 отщепляются. ГТФ, связанный с IF-2, гидролизуется до ГДФ и Фн (неорганический фосфат). Комплекс IF-2 + ГДФ диссоциирует от 70S рибосомы, связанной в P-сайте с формилметионил-тРНК.
9В роли старт-кодона у прокариот может выступать не только AUG (83% генов), но и GUG (14% генов; кодирует валин), и UUG (3% генов; кодирует лейцин).
10Формилметионин — аминокислота метионин, связанная с формильной группой (—
CHO).
122 |
Глава 6 |
Трансляция |
Рис. 81. Инициация трансляции у прокариот.
Рис. 82. Узнавание последовательности Шайна-Дальгарно у E. coli.
Инициация у эукариот 123
6.6 Инициация у эукариот
Инициация у про- и эукариот схожа, однако есть и различия:
1.Эукариотическая 40S субъединица рибосом связывается с факторами eIF1A и eIF-3. Затем к 40S частице присоединяется фактор eIF-2 + ГТФ + меттРНК. Формируется преинициационный комплекс 43S.
2.43S комплекс связывается с факторами eIF-4E, eIF-4G, eIF-4A, eIF-4B и мРНК в области 5’-кэпа. У эукариотических мРНК отсутствует последовательность Шайна-Дальгарно, поэтому у них существует сканирующий механизм обнаружения старт-кодона AUG11. eIF-40S комплекс движется по
цепи мРНК до т.н. последовательности Ко́зака (–3–ACCAUGG–+4). Рибо-
сома может пропустить первый встретившийся ей кодон AUG, если он не образует последовательность Козака. В конце данного этапа формируется 48S комплекс.
3.Происходит гидролиз ГТФ, факторы инициации отщепляются, присоединяется 60S субъединица и формируется 80S инициационный комплекс.
6.7Элонгация у прокариот
Элонгация — это вторая стадия трансляции, на которой синтезируется сама полипептидная цепь. Элонгация включает три важнейших этапа:
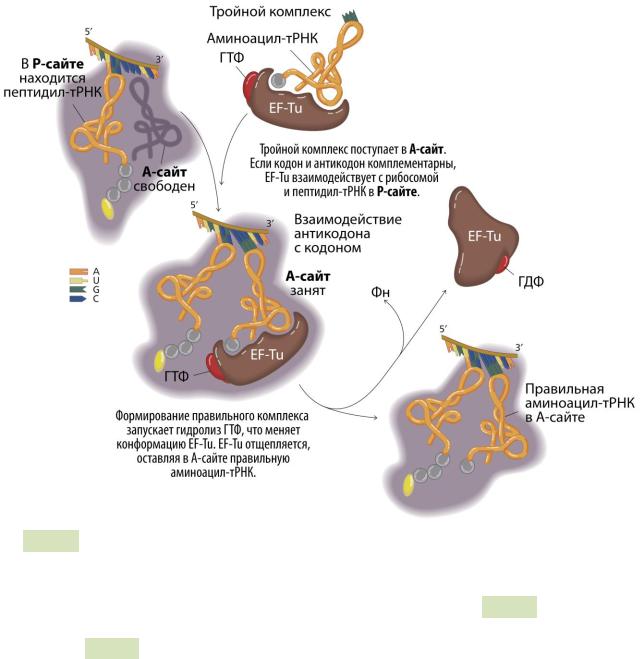
1.Декодирование (связывание аминоацил-тРНК, несущей аминокислоту) (см. Рис. 83 ▼). После инициации в P-сайте рибосомы уже находится инициирующая формилметионил-тРНК. В А-сайте происходит связывание второй аминоацил-тРНК, комплементарной своим антикодоновым участком кодону в мРНК. В этом процессе участвует фактор EF-Tu, направляющий аминоацил-тРНК в А-сайт. Если антикодон на аминоацил-тРНК оказывается комплементарен кодону на мРНК, происходит стабилизация комплекса EF-Tu + ГТФ + аат-тРНК и гидролиз ГТФ до ГДФ и Фн. EF-Tu + ГДФ диссоциирует от рибосомы, связывается с фактором EF-Ts. ГДФ обменивается на ГТФ и EF-Ts отщепляется от EF-Tu.
11Кодон AUG у эукариот кодирует аминокислоту метионин (не формилметионин, как
упрокариот).
124 |
Глава 6 |
Трансляция |
Рис. 83. Элонгация трансляции у прокариот: декодирование и «вставка» аминоа-
цил-тРНК в А-сайт.
2.Транспептидация (образование пептидной связи) (см. Рис. 84 ▼). Происходит образование пептидной связи по механизму нуклеофильной атаки (см. Рис. 85 ▼). Реакция катализируется пептидилтрансферазным центром большой субъединицы рибосом (катализируется не белком, а молекулой РНК в составе рибосом). Растущая полипептидная цепь оказывается связанной с тРНК в А-сайте. В Р-сайте находится уже свободная тРНК.
3.Транслокация (перемещение мРНК относительно рибосомы на один кодон). Третий фактор элонгации EF-G + ГТФ связывается с рибосомой. Происходит гидролиз ГТФ до ГДФ и Фн. EF-G + ГДФ отщепляется, происходит транслокация мРНК относительно рибосомы. То есть именно цепочка мРНК движется относительно неё, а не наоборот. Свободная тРНК перемещается из P-сайта в E-сайт. Пептидил-тРНК переносится из A-сайта в P-
сайт. А-сайт остаётся свободным.
Элонгация — циклический процесс. Он повторяется ровно столько раз, сколько кодонов находится между старт- и стоп-кодоном в цепи мРНК.