

Вопрос №77
Специфический обмен аминокислот: глицина, серина. Роль Н4-фолата и его производных
в обмене аминокислот и нуклеотидов.
Серин - заменимая аминокислота, синтезируется из промежуточного продукта гликолиза - 3-фосфоглицерата, а аминогруппу получает от глутаминовой кислоты.
Г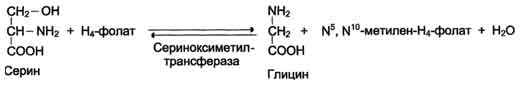 лицин
- также заменимая аминокислота, основным
источником которой служит серии. Реакцию
синтеза глицина из серина катализирует
фермент серин-оксиметилтрансфераза,
кофермен-том которой является Н4-фолат.
лицин
- также заменимая аминокислота, основным
источником которой служит серии. Реакцию
синтеза глицина из серина катализирует
фермент серин-оксиметилтрансфераза,
кофермен-том которой является Н4-фолат.
Р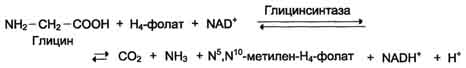 еакция
превращения серина в глицин легко
обратима. Основной путь катаболизма
глицина у человека и других позвоночных
также связан с использованием Н4-фолата.
еакция
превращения серина в глицин легко
обратима. Основной путь катаболизма
глицина у человека и других позвоночных
также связан с использованием Н4-фолата.
Эта реакция обратима и катализируется глицинсинтазой - ферментным комплексом, похожим на пируватдегидрогеназный комплекс, и локализованным в митохондриях клеток печени. По последним данным глицинрасщепляющая ферментная система несколько отличается от глицинсинтазы и содержит 4 белка: Р-белок, включающий кофермент ПФ, Н-белок, содержащий липоевую кислоту, Т-белок с коферментом Н4-фолат, L-белок, являющийся дигидролипоилдегидрогеназой с коферментом NAD+.
1. Пути метаболизма
серина и глицина. О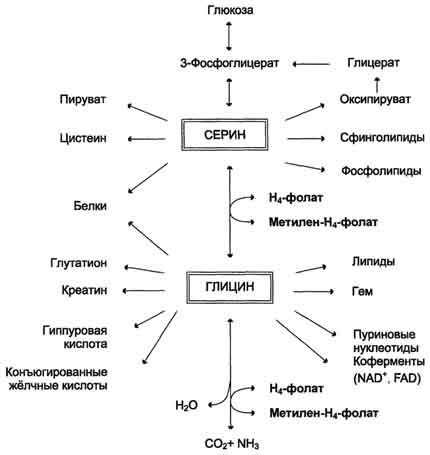 бе
аминокислоты необходимы не только для
синтеза белков и глюкозы (при её недостатке
в клетках), но и нукле-отидов, коферментов,
тема, сложных липидов, креатина и других
соединений. Многие из этих реакций
представлены в соответствующих разделах
учебника.
бе
аминокислоты необходимы не только для
синтеза белков и глюкозы (при её недостатке
в клетках), но и нукле-отидов, коферментов,
тема, сложных липидов, креатина и других
соединений. Многие из этих реакций
представлены в соответствующих разделах
учебника.
Роль фолиевой кислоты в обмене
В превращениях серина и глицина главную роль играют ферменты, коферментами которых служат производные фолиевой кислоты. Этот витамин широко распространён в животных и растительных пищевых продуктах. Молекула фолиевой кислоты (фолата) состоит из 3 частей: птеринового производного, парааминобензойной и глутаминовой кислот.
Коферментную функцию выполняет восстановленная форма фолата - тетрагидрофолиевая кислота (ТГФК или Н4-фолат).
Ф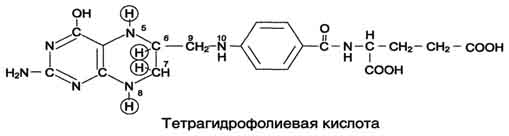 олиевая
кислота в печени превращается в Н4-фолат
в несколько стадий с участием ферментов
фолатредуктазы и дигидрофолатредуктазы,
коферментом которых служит NADPH.
олиевая
кислота в печени превращается в Н4-фолат
в несколько стадий с участием ферментов
фолатредуктазы и дигидрофолатредуктазы,
коферментом которых служит NADPH.
Н4-фолат - акцептор β-углеродного атома серина. При этом образуется метиленовый мостик между атомами азота в молекуле Н4-фолата в положениях 5 и 10, образуя метилен-Н4-фолат
Дефицита фолиевой - мегалобластная анемия-уменьшением количества эритроцитов, снижением гемоглобина, увеличение размера эритроцитов. Причина - нарушение синтеза ДНК и РНК из-за недостатка предшественников - тимидиловой кислоты и пуриновых нуклеотидов вследствие дефицита производных Н4-фолата.
Вопрос №78
Основные пути превращения метионина в организме;
значение реакций трансметилирования. Синтез креатина.
М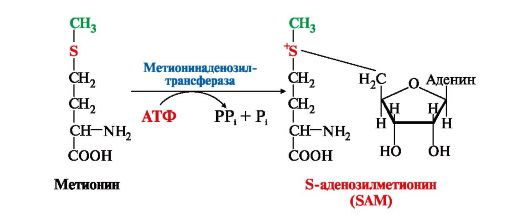 етионин
- незаменимая аминокислота, необходимая
для синтеза белков. Мет-тРНКмет участвует
в инициации процесса трансляции каждого
белка. Как и многие другие аминокислоты,
метионин подвергается транс- и
дезаминированию. Особая роль метионина
заключается в том, что метильная группа
этой аминокислоты используется для
синтеза целого ряда соединений в реакциях
трансметилирования. Основным донором
метильной группы является S-аденозилметионин
(SAM) - активная форма метионина, который
присутствует во всех типах клеток и
синтезируется из метионина и АТФ под
действием фермента метионин-аденозилтрансферазы.
етионин
- незаменимая аминокислота, необходимая
для синтеза белков. Мет-тРНКмет участвует
в инициации процесса трансляции каждого
белка. Как и многие другие аминокислоты,
метионин подвергается транс- и
дезаминированию. Особая роль метионина
заключается в том, что метильная группа
этой аминокислоты используется для
синтеза целого ряда соединений в реакциях
трансметилирования. Основным донором
метильной группы является S-аденозилметионин
(SAM) - активная форма метионина, который
присутствует во всех типах клеток и
синтезируется из метионина и АТФ под
действием фермента метионин-аденозилтрансферазы.
Структура -S+-CH3 в SAM является нестабильной, метильная группа легко отщепляется, что определяет высокую способность ее к переносу на другие соединения в реакциях трансметилирования.
В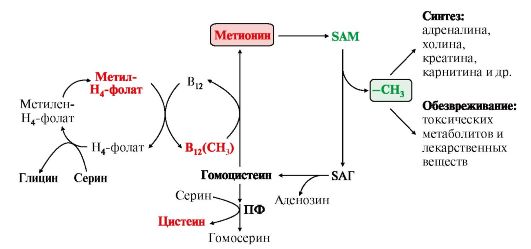 реакциях трансметилирования SAM
превращается в S-аденозилгомо- цистеин
(SAr), который гидролитически расщепляется
с образованием аденозина и гомоцистеина.
Последний может снова превращаться в
метионин с участием метил-Н4-фолата и
витамина В12. Регенерация метионина
тесно связана с обменом серина и глицина
и взаимопревращениями производных
Н4-фолата.
реакциях трансметилирования SAM
превращается в S-аденозилгомо- цистеин
(SAr), который гидролитически расщепляется
с образованием аденозина и гомоцистеина.
Последний может снова превращаться в
метионин с участием метил-Н4-фолата и
витамина В12. Регенерация метионина
тесно связана с обменом серина и глицина
и взаимопревращениями производных
Н4-фолата.
Метионин и серин необходимы для синтеза условно заменимой аминокислоты цистеина, причем в этом процессе метионин является донором атома серы. Цистеин образуется непосредственно из гомоцистеина в ходе двух реакций, которые происходят с участием пиридоксальфосфата. Генетический дефект этих ферментов приводит к нарушению использования гомоцистеина в организме и превращению его в гомоцистин, который может накапливаться в крови и тканях, выделяться с мочой, вызывая гомоцистинурию. Заболевание сопровождается эктопией хрусталика глаза, катарактой, остеопорозом, умственной отсталостью.
С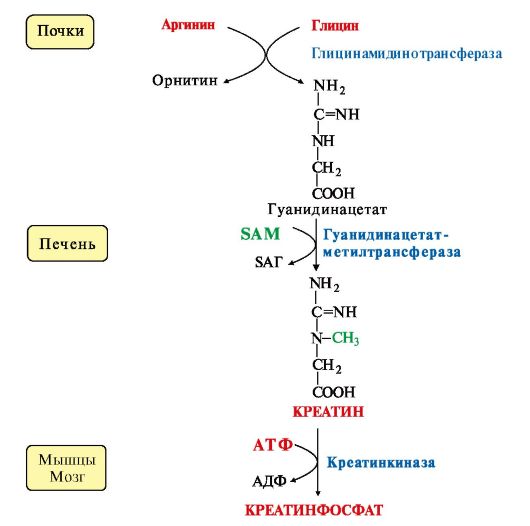 интез
креатина
происходит с использованием трех
аминокислот: аргинина, глицина и
метионина. Процесс начинается в почках,
в реакцию вступают аргинин и глицин.
Образующийся гуанидинацетат поступает
в печень, где подвергается метилированию
с участием SAM и превращается в креатин.
Из печени креатин транспортируется в
мышцы и головной мозг. Креатин в клетках
превращается в креатинфосфат -
макроэргическое соединение, являющееся
резервной формой энергии в мышечной и
нервной тканях. Содержание креатинфосфата
в покоящейся мышце в восемь раз выше,
чем АТФ. Эту реакцию катализирует фермент
креатинкиназа.
интез
креатина
происходит с использованием трех
аминокислот: аргинина, глицина и
метионина. Процесс начинается в почках,
в реакцию вступают аргинин и глицин.
Образующийся гуанидинацетат поступает
в печень, где подвергается метилированию
с участием SAM и превращается в креатин.
Из печени креатин транспортируется в
мышцы и головной мозг. Креатин в клетках
превращается в креатинфосфат -
макроэргическое соединение, являющееся
резервной формой энергии в мышечной и
нервной тканях. Содержание креатинфосфата
в покоящейся мышце в восемь раз выше,
чем АТФ. Эту реакцию катализирует фермент
креатинкиназа.
Креатинфосфат играет важную роль в обеспечении работающей мышцы энергией в начальный период физической работы. В работающей мышце концентрация АТФ некоторое время остается постоянной, а концентрация креатинфосфата быстро снижается. Часть образовавшегося креатина и креатинфосфата с постоянной скоростью превращается в креатинин, который выводится с мочой (норма - 1-2 г/сут, или 8,8-17,6 ммоль/л). При уменьшении массы мышц вследствие длительного отрицательного азотистого баланса, при состояниях, ведущих к атрофии мышц, выделение креатинина снижается (голодание, острые инфекции, сахарный диабет, гипертиреоз).