

Вопрос № 43
ФАД-зависимые дегидрогеназы и их субстраты. Строение окисленной и восстановленной форм ФАД. Путь электронов в дыхательной цепи от субстратов, окисляемых ФАД-зависимыми дегидрогеназами. Коэффициент Р/О.
Д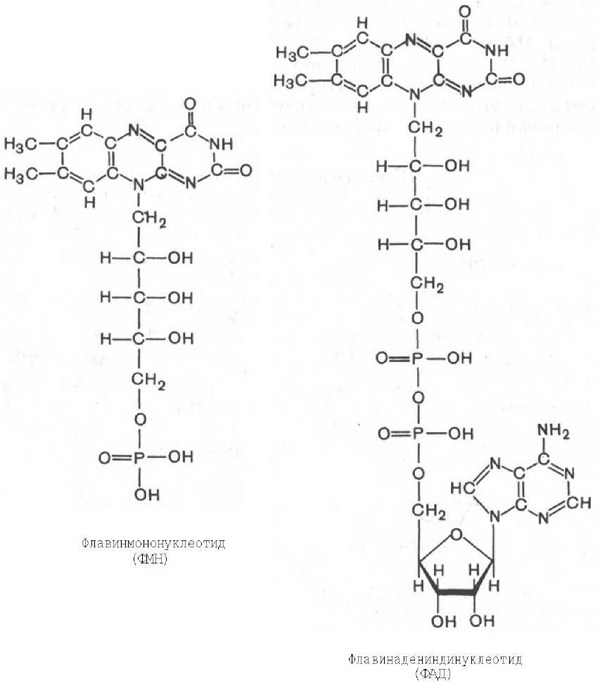 егидрогеназы
окисляют субстрат путем отнятия
водорода. Со-ферментная часть дегидрогеназ
представлена флавиновыми нуклеотидами
(ФМН, ФАД). Флавиновые дегидрогеназы
могут быть и анаэробными, и аэробными.
Окисление с участием флавиновых
дегидрогеназ аэробного типа, как
правило, ведет к образованию пероксида
водорода. Эти дегидрогеназы могут быть
названы и оксидазами из-за их взаимодействия
с кислородом. Флавиновые дегидрогеназы
отличются большим разнообразием
окислительно-восстановительного
потенциала, они нередко содержат металлы
- медь, молибден, железо.
егидрогеназы
окисляют субстрат путем отнятия
водорода. Со-ферментная часть дегидрогеназ
представлена флавиновыми нуклеотидами
(ФМН, ФАД). Флавиновые дегидрогеназы
могут быть и анаэробными, и аэробными.
Окисление с участием флавиновых
дегидрогеназ аэробного типа, как
правило, ведет к образованию пероксида
водорода. Эти дегидрогеназы могут быть
названы и оксидазами из-за их взаимодействия
с кислородом. Флавиновые дегидрогеназы
отличются большим разнообразием
окислительно-восстановительного
потенциала, они нередко содержат металлы
- медь, молибден, железо.
Ф АД-зависимые
субстраты тканевого дыхания – сукцинат,
глицерол-3-фосфат, ацил~КоА и некоторые
другие. Водород от ФАД-зависимых
субстратов передаётся на II-й комплекс
дыхательной цепи.
АД-зависимые
субстраты тканевого дыхания – сукцинат,
глицерол-3-фосфат, ацил~КоА и некоторые
другие. Водород от ФАД-зависимых
субстратов передаётся на II-й комплекс
дыхательной цепи.
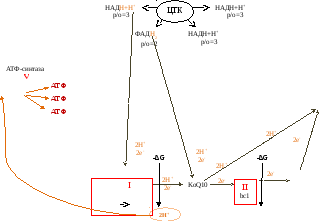
Механизм переноса электронов и образования воды
Электроны могут поступать в ЦПЭ от НАДН+Н+ и ФАД∙Н2.
От ФАД∙Н2 – на Q10.
Он становится QH2.
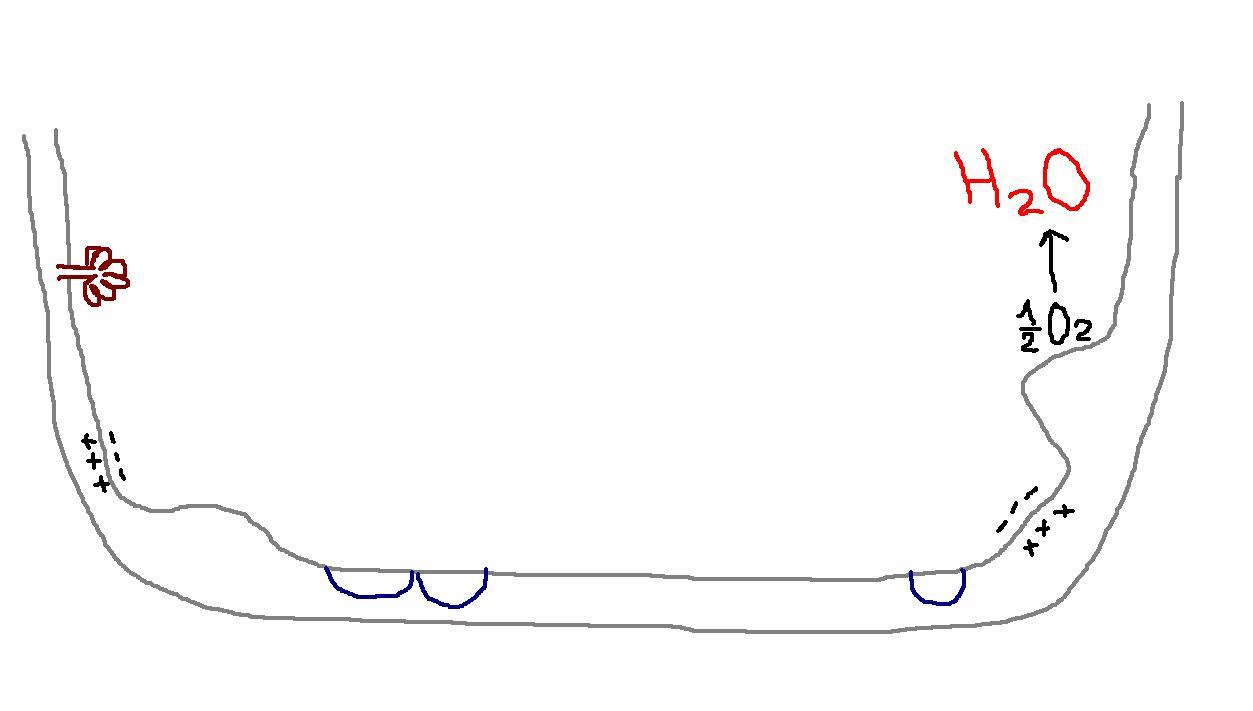
От QH2 по системе цитохромов передаются только электроны, а протоны поступают матрикс и идут на восстановление воды.
На этом тканевое дыхание заканчивается.
НАДН-ДГ
ФМН FeS
Роль передачи электронов в синтезе АТФ
В начало цепи поступают электронов, богатые энергией.
В процессе переноса из-за разности ОВ-потенциала они теряют эту энергию на каждом участке (около 200 кДж). Этого хватило бы на синтез 4 молекул АТФ, но синтезируется всего 3. Остальная энергия рассеивается для поддержания температуры тела.
Больше всего свободной энергии выделяется в трех участках. Они называются участками сопряженного синтеза АТФ (I, III, IV ФК).
Свободная энергия Гиббса, которая выделяется при переносе электронов в трех участках. Направлена на выталкивание протонов из матрикса в межмембранное пространство.
В результате наружная сторона внутренне мембраны заряжается положительно, а внутренняя – отрицательно.
Создается трансмембранный электрохимический протонный градиент. Он необходим для последующего синтеза АТФ.
Протоны по своему градиенту стремятся вернуться назад в матрикс. Но внутренняя мембрана для них не проницаема, и они возвращаются в матрикс через специфический F1-комплекс (АТФ-синтаза, V ФК).
F1-комплекс состоит из двух частей:
F0 – канал в мембране (из 6 субъединиц)
F1 = фермент АТФ-синтаза – синтез АТФ при прохождении протонов (6 субъединиц).
Каждая пара протонов, проходящая через F1-комплекс, активирует АТФ-синтазу, которая синтезирует 1 молекулу АТФ.
Таким образом, если е- и Н+ поступают в начало цепи (от ФАД∙Н2), то в процессе сопряженного синтеза образуется 2 АТФ.
Следовательно, за счет 4 реакций ЦТК синтезируется 11 АТФ (3х3+1х2).
Коэффициент фосфорилирования (р/о) - количество АТФ, которое образуется при переносе электронов к одному атому кислорода. Он может быть равен 3, 2, 1.
Вопрос №44
Современные представления о механизме окислительного фосфорилирования. Хемоосмотическая теория Митчелла.
Теорию создания протонного градиента и его роль в синтезе АТФ создал Митчелл (хеми-осмотическая теория):
«Компоненты дыхательной цепи расположены строго определенным образом так, что при передаче пары электронов по ЦПЭ энергия, вылеляющаяся в трех участках, направлена на выталкивание протонов в межмембранное пространство. В результате создается трансмембранный электрохимический протонный градиент – движущая сила синтеза АТФ» (путем окислительного фосфорилирования).