

Вопрос №33
Биосинтез ДНК. Механизм воспроизведения (репликация).
Матрица – двуцепочечная молекула ДНК.
Субстрат (из чего синтезируется продукт) – дНТФ (дезоксирибонуклеозидтрифосфаты): дАТФ, дГТФ...
Продукт – две дочерние ДНК, комплементарные друг другу (но не идентичные).
Источник энергии – дНТФ (дезоксирибонуклеозидтрифосфаты): дАТФ, дГТФ, дЦТФ, дТТФ.
Ферменты – топоизомераза I,II, ДНК-полимераза, ДНК-лигаза.
Локализация в клетке – ядро (S-фаза клеточного цикла).
Стадии: 1)Образование репликативной вилки, 2) Синтез дочерних цепей ДНК, 3)Удаление праймеров
Основной механизм – полуконсервативный, т.е. двойная спираль раскручивается, цепи расходятся и каждая затем достраивается до двуцепочечной по принципу комплементарности. Каждая новая двуцепочечная молекула ДНК = 1 цепь старой + 1 цепь дочерней ДНК.
1. Образование репликативной вилки: Топоизомеразы I (II) находят на двуцепочечной молекуле ДНК определенную последовательность – точку начала репликации. Затем разрывают или одну цепь (топоизомераза I), или обе (топоизомераза II) и встраиваются в них.
Происходит разрыв двуцепочечной молекулы с образованием репликативного «глаза» = участок инициации репликации = original.
Т.к. молекула ДНК содержит около 150∙106 пар нуклеотидов, а репликация идет со скоростью 50 нуклеотидов/мин, то репликация занимала был 800 часов, а она идет 9.
Следовательно, на молекуле ДНК несколько участков начала репликации, и репликация идет в двух направлениях.
В репликативном глазе можно выделить 2 репликативные вилки.
Специфический фермент ДНК-геликаза (активность зависит от АТФ) встраивается между цепями и разъединяет их.
Вдоль цепи расположены ДНК-раскручивающие белки = ДНК-дестабилизирующие белки = SSB-белки, которые не позволяют цепям ДНК образовать двуцепочную структуру, удерживают их в раскрученном виде, облегчают репликацию.
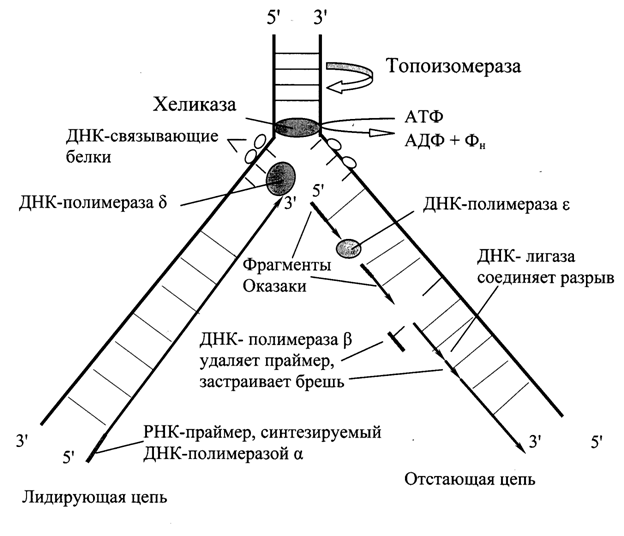
2.Синтез дочерних цепей.
ДНК-полимеразы:
Осуществляют синтез в направлении 5’→3’ дочерней цепи
Не могут начинать синтез, а могут только достраивать.
Следовательно, для синтеза дочерних цепей необходима «затравка» = праймер.
Праймер строится из нуклеотидов, принадлежащих РНК (чтобы отличить его от дочерних цепей). Для этого используется фермент РНК-праймаза.
После этого ДНК-полимераза-δ синтезирует дочернюю цепь быстро и непрерывно, т.к. направление синтеза совпадает с направлением репликации.
Вторая дочерняя цепь синтезируется медленнее и фрагментами (фрагменты Оказаки), т.к. по принципу антипараллельности ДНК-полимераза достраивала бы в направлении 3’→ 5’, чего не может быть. Если бы мы строили в нужном направлении, то синтез дочерних цепей не совпадал бы с общим направлением репликации. Поэтому отстающая цепь синтезируется фрагментами.
Для этого на материнской цепи строится несколько РНК-праймеров (около 200 нуклеотидов).
ДНК-полимераза-ε узнает 3’-конец праймера и достраивает его в нужном 5’→3’ направлении до предыдущего праймера.
Далее ДНК- полимераза-ε двигается в направлении репликации и находит 3’-конец следующего праймера и достраивает его (фрагменты Оказаки). И так далее.
РНК-аза (эндонуклеаза) вырезает праймеры. ДНК-полимераза-β достраивает недостающие участки. Одноцепочечные разрывы соединяют ДНК-лигазы.
Пострепликативные модификации ДНК: После синтеза ДНК материнская цепь отличается от дочерней, т.к. в ней есть метилированный аденин в последовательности Г-А-Т-Ц. Это позволяет ферментам системы репарации исправлять ошибки, которые могут возникнуть в дочерних цепях. После этого и дочерняя цепь метилируется по аденину в последовательности Г-А-Т-Ц.