

Тема 9.11. Обмен фенилаланина, тирозина и гистидина в разных тканях
|
|
1. Фенилаланин - незаменимая аминокислота, так как в клетках животных не синтезируется ароматическое кольцо. Основная часть поступающего с пищей фенилаланина используется в синтезе тканевых белков, превращение остальной части начинается с его гидроксилирования, в результате чего образуется тирозин. Реакция эта катализируется специфической монооксигеназой - фенилаланингидроксилазой, коферментом которой является тетрагидробиоптерин (Н4-БП) (рис. 9.22).
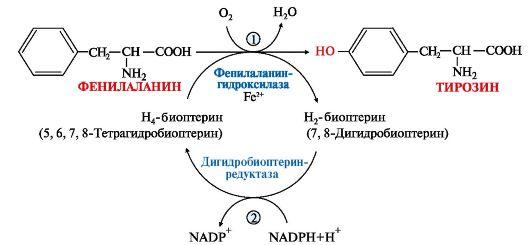 Рис.
9.22. Реакция гидроксилирования фенилаланина
и регенерация тетрагидробиоптерина
(Н4БП):
Рис.
9.22. Реакция гидроксилирования фенилаланина
и регенерация тетрагидробиоптерина
(Н4БП):
1. Реакцию катализирует фенилаланингидроксилаза (1), коферментом которой является Н4БП. Кофактором являются ионы Fe2+. Н4БП в результате реакции окисляется в дигидробиоптерин (Н2БП).
2. Регенерация дигидробиоптерина (2) происходит при участии дигидробиоптеринредуктазы с использованием NADPH.
2. Тирозин - условно заменимая аминокислота. Синтезируется только из фенилаланина.
Катаболизм Фен и Тир происходит в печени. В результате ряда реакций образуется фумарат и ацетоацетат (рис. 9.23, А). Фумарат используется для синтеза глюкозы (глюконеогенез) или окисляется до СО2 и Н2О. Ацетоацетат - кетоновое тело, которое окисляется в тканях с выделением энергии. Таким образом, Фен и Тир относятся к смешанным (гликокетогенным) аминокислотам по использованию безазотистого остатка.
Превращение промежуточного продукта катаболизма Тир - гомогентизиновой кислоты - в фумарилацетоацетат сопровождается расщеплением ароматического кольца.
 Процессы
расщепления ароматических колец в
биологических системах катализируются
ферментами диоксигеназами. Для
катализа диоксигеназам необходимы кофакторы
- Fe2+ или гем (для
некоторых - Сu+), а также - витамин
С.
Процессы
расщепления ароматических колец в
биологических системах катализируются
ферментами диоксигеназами. Для
катализа диоксигеназам необходимы кофакторы
- Fe2+ или гем (для
некоторых - Сu+), а также - витамин
С.
3. В мозговом веществе надпочечников и нервной ткани из тирозина синтезируются катехоламины (дофамин, норадреналин, адреналин) (рис. 9.23, В).
|
|
Тирозин под действием специфической монооксигеназы - тирозингидроксилазы превращается в ДОФА. Для протекания реакции необходимы Н4БП, О2 и Fe2+ (реакция аналогична гидроксилированию фенилаланина, см. рис. 9.22). Тирозингидроксилаза найдена только в надпочечниках и катехоламинэргических нейронах (преимущественно в их нервных окончаниях). Этот фермент является регуляторным и определяет скорость синтеза катехоламинов. Одна из функций последних - регуляция деятельности сердечнососудистой системы.
Активность тирозингидроксилазы значительно изменяется в результате:
• аллостерической регуляции по принципу ретроингибирования норадреналином;
• фосфорилирования с участием цАМР - зависимой протеинкиназы, при этом снижается Км для кофермента Н4БП и сродство фермента к норадреналину, в результате чего происходит активация тирозингидроксилазы;
• индукции синтеза фермента кортизолом.
Катехоламины выполняют очень важные функции в организме. Дофамин является медиатором среднего отдела мозга. Норадреналин - тормозный медиатор синаптической нервной системы и разных отделов головного мозга, но может выполнять функцию возбуждающего медиатора в гипоталамусе. Адреналин - гормон интенсивной физической работы, который синтезируется при стрессе и регулирует основной обмен, а также усиливает сокращение сердечной мышцы.
В щитовидной железе тирозин используется для синтеза гормонов иодтиронинов (тироксина и трииодтиронина) (рис. 9.23, Г). Подробно их функции и синтез рассматриваются в модуле 11.
В меланоцитах - пигментных клетках кожи, сетчатки глаз тирозин является предшественником пигментов меланинов (см. рис. 9.23, Б).
4. Частично заменимая аминокислота гистидин синтезируется из глутамата в сложном процессе, поскольку образование гетероциклического радикала в клетках человека и млекопитающих сопряжено с большими трудностями.
|
|
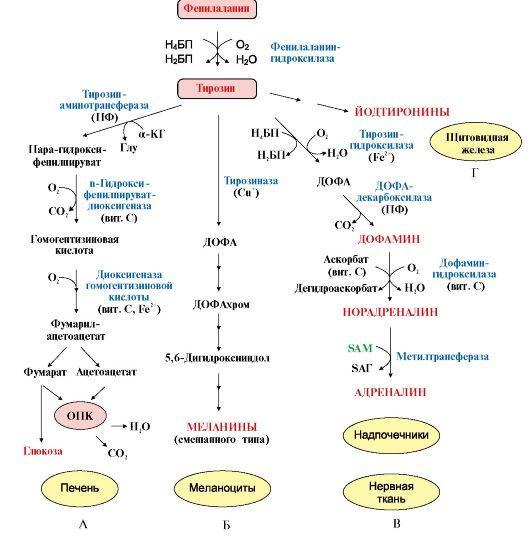 Рис.
9.23. Пути превращения фенилаланина и
тирозина в разных тканях:
Рис.
9.23. Пути превращения фенилаланина и
тирозина в разных тканях:
А - катаболизм фенилаланина и тирозина в печени; Б - синтез меланинов в меланоцитах; В - синтез катехоламинов в надпочечниках и нервной ткани; Г - синтез иодтиронинов в щитовидной железе
Обмен гистидина включает синтез гистамина в соединительной ткани, а также путь катаболизма, который происходит в печени и, частично, в коже человека.
В печени и коже дезаминирование гистидина катализирует фермент гистидаза. Образующийся уроканат только в печени способен превращаться через ряд стадий в глутамат. Наследственный дефект гистидазы вызывает
накопление гистидина в организме и развитие гистидинемии, которая проявляется задержкой в умственном и физическом развитии детей. Ферменты гистидаза и уроканиназа (рис. 9.24) являются гепатоспецифическими, поэтому их определение используется в клинике для диагностики поражений печени.
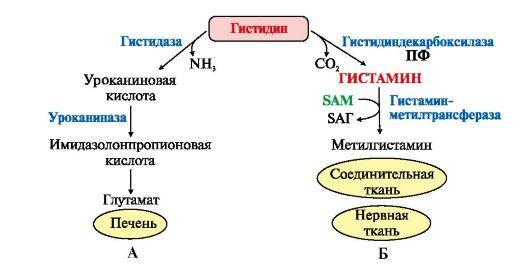 Рис.
9.24. Схема обмена гистидина в разных
тканях:
Рис.
9.24. Схема обмена гистидина в разных
тканях:
А - катаболизм гистидина в печени; Б - синтез и инактивация гистамина
7. Гистамин синтезируется путем декарбоксилирования гистидина в тучных клетках соединительной ткани, образует комплекс с белками и сохраняется в секреторных гранулах. Выделяется в кровь при повреждении ткани (удар, ожог, воздействие эндо- и экзогенных веществ). Гистамин выполняет следующие функции:
• стимулирует секрецию желудочного сока, слюны (пищеварительный гормон);
• обеспечивает воспалительную реакцию - расширение сосудов, покраснение кожи, отечность ткани;
• обеспечивает аллергическую реакцию;
• повышает проницаемость капилляров, вызывает отеки, снижает артериальное давление (но увеличивает внутричерепное давление, вызывает головную боль);
• сокращает гладкую мускулатуру легких, вызывает удушье;
|
|
• выполняет роль нейромедиатора.