

Билет 2.
Генотипическая и фенотипическая изменчивость бактерий. Регуляция экспрессии генов.



В 1961 году Жакоб и Моно установили, что у бактерий под контролем гена-регулятора синтезируется белок-репрессор, который регулирует активность других генов. Белок-репрессор связывается с геном-оператором и блокирует его, вследствие чего транскрипция (считывание информации) становится невозможной и клетка не может синтезировать соответствующие белки. Но если в клетку проникает какой-либо субстрат, для расщепления которого необходимо синтезировать ферменты, то происходит следующее: субстрат присоединяется к репрессору и лишает его способности блокировать ген-оператор.
К ДНК присоединяются РНК-синтезирующие ферменты и начинается транскрипция генов. Синтез белка-фермента прекращается, если субстрат полностью расщепляется, белок-репрессор освобождается и снова блокирует гены-операторы. Это пример регуляции по типу индукции. Также существует регуляция по типу репрессии: когда конечные продукты биохимической реакции, соединяясь с неактивным белком-репрессором, образуют комплекс, блокирующий работу гена-оператора.
Антитерминация – заключается в том, что происходит игнорирование терминальных кодонов, процесс экспрессии продолжается.
Изучение регуляции генной активности у прокариот привело французских микробиологов Ф. Жакоба и Ж. Моно к созданию (1961) оперонной модели регуляции транскрипции. Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. Например, это могут быть гены, которые детерминируют синтез ферментов, участвующих в метаболизме какого-либо вещества или в синтезе какого-то компонента клетки. Оперонная модель регуляции экспрессии генов предполагает наличие единой системы регуляции у таких объединенных в один оперон структурных генов, имеющих общий промотор и оператор.
Особенностью прокариот является транскрибирование мРНК со всех структурных генов оперона в виде одного полицистронного транскрипта, с которого в дальнейшем синтезируются отдельные пептиды.
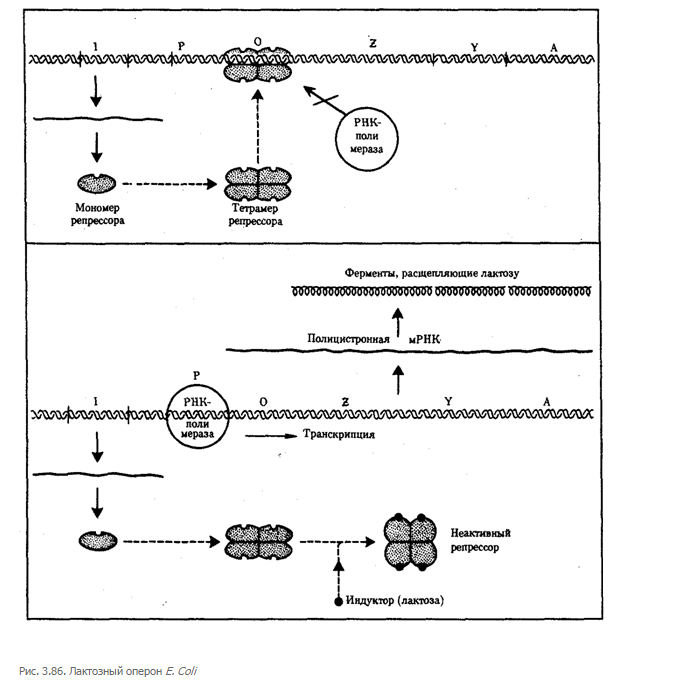
Примером участия генетических и негенетических факторов в регуляции экспрессии генов у прокариот может служить функционирование лактозного оперона у кишечной палочки Е. colt (рис. 3.86). При отсутствии в среде, на которой выращиваются бактерии, сахара лактозы активный белок-репрессор, синтезируемый геном-регулятором (I), взаимодействует с оператором (О), препятствуя соединению РНК-полимеразы с промотором (Р) и транскрипции структурных генов Z, Y, А. Появление в среде лактозы инактивирует репрессор, он не соединяется с оператором, РНК-полимераза взаимодействует с промотором и осуществляет транскрипцию полицистронной мРНК. Последняя обеспечивает синтез сразу всех ферментов, участвующих в метаболизме лактозы. Уменьшение содержания лактозы в результате ее ферментативного расщепления приводит к восстановлению способности репрессора соединяться с оператором и прекращению транскрипции генов Z, Y, А.
Таким образом, регуляция экспрессии генов, организованных у прокариот в опероны, является координированной. Синтез полицистронной мРНК обеспечивает одинаковый уровень синтеза всех ферментов, участвующих в биохимическом процессе.
Патогенность и вирулентность бактерий. Факторы патогенности.




