

16.2.2. Стационарные уровни концентрации биомассы и продукта ферментации
На основе выражения (16.28) для стационарных уровней концентраций субстрата с использованием уравнения (16.12) могут быть найдены стационарные уровни концентрации биомассы:
![]() (16.41)
(16.41)
или
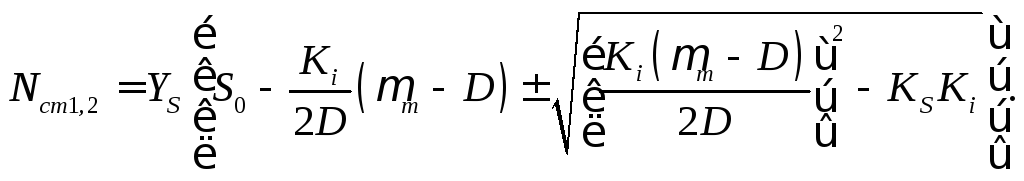
(16.42)
Рассмотрим зависимости стационарной концентрации биомассы от параметров процесса с использованием известных приближений. При D >Dc стационарного состояния по концентрации биомассы не существует. Если D=Dc, то имеет место одно стационарное состояние, в котором
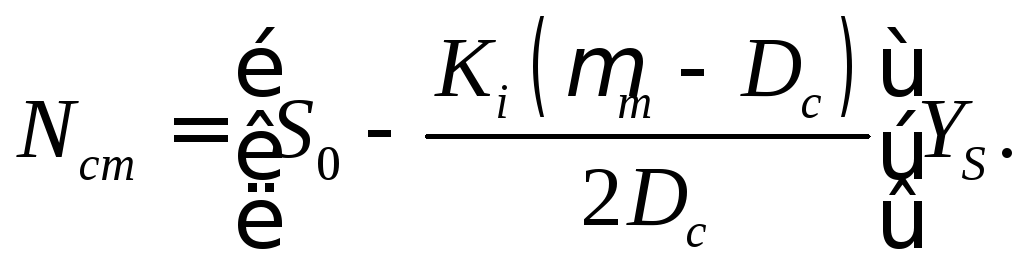 (16.43)
(16.43)
В этом режиме стационарная концентрация биомассы линейно зависит от начальной концентрации вводимого субстрата. Если в уравнение (16.43) подставить значение Dс в соответствии с уравнением (16.32), то будем иметь
![]() (16.44)
(16.44)
Стационарная концентрация биомассы в точке D=Dc уменьшается с ростом константы сродства субстрата и константы ингибирования процесса избытком субстрата. При этом стационарная концентрация биомассы не зависит от максимальной скорости роста микроорганизма.
При уменьшении скорости разбавления в режиме D<Dc концентрация биомассы может иметь одно из двух стационарных значений, которые даются иррациональными функциями (16.42). Однако в режиме D<<Dc при учете соотношений (16.39)— (16.40) имеем
 (16.45)
(16.45)
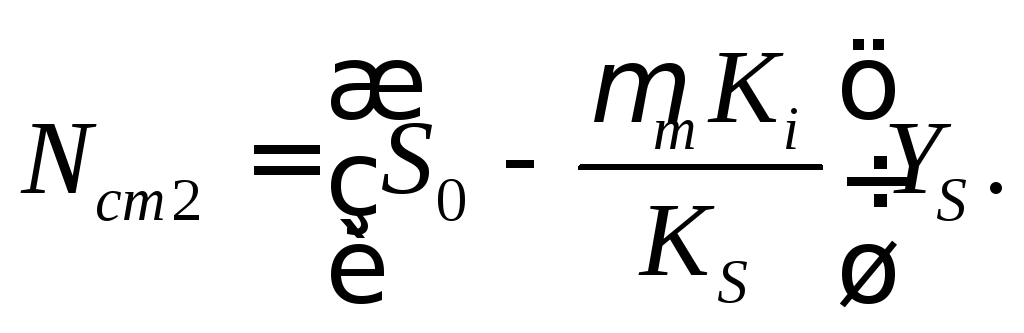 (16.46)
(16.46)
Первый
стационарный уровень тем выше, чем выше
максимальная удельная скорость роста
культуры и чем ниже скорость
разбавления. Для второго уровня характерно
обратное: чем выше скорость разбавления
и чем ниже максимальная скорость роста
микроорганизма, тем выше значение
стационарной концентрации биомассы.
При
уменьшении скорости разбавления D→0
Nст
1→YSS0,
Nст
2→
-∞(при
![]() стационарная
концентрация биомассы равна нулю) (рис.
16.7). Концентрация продукта ферментационного
превращения в стационарном режиме
линейно связана с концентрацией биомассы
(16.14):
стационарная
концентрация биомассы равна нулю) (рис.
16.7). Концентрация продукта ферментационного
превращения в стационарном режиме
линейно связана с концентрацией биомассы
(16.14):
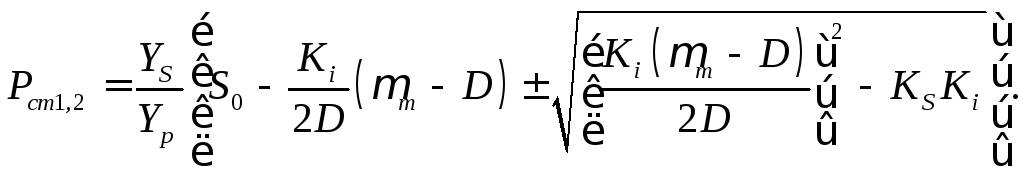 (16.47)
(16.47)
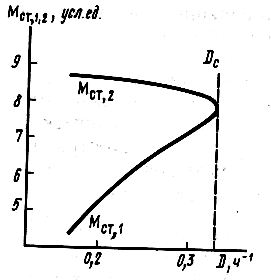
|
Рис.16.7. Стационарное состояние в хемостате для культуры ингибируемой избытком субстрата |
В режиме D=Dc имеется одно стационарное состояние концентрации продукта:
![]() (16.48)
(16.48)
При D<<Dc наблюдается два стационарных состояния с концентрациями:
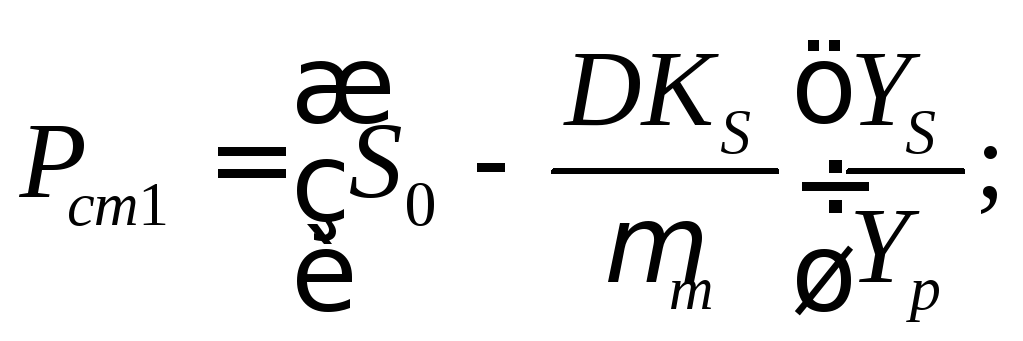 (16.49)
(16.49)
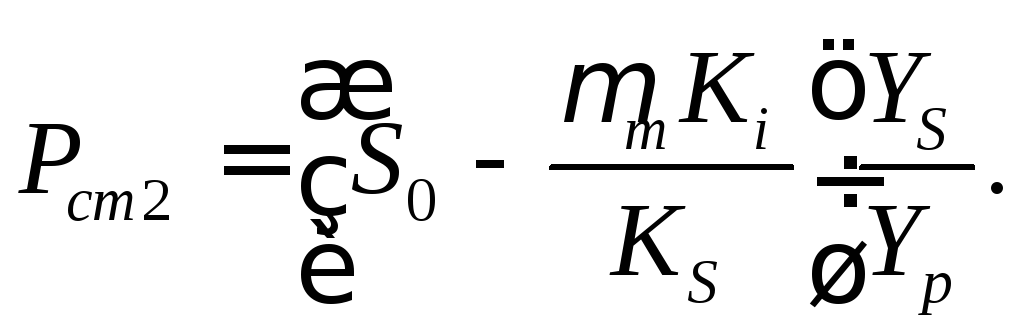 (16.50)
(16.50)
Таким образом, наиболее важной особенностью развития процесса в хемостате для микроорганизма с ингибированием роста избытком субстрата является возможность образовывать два различных стационарных состояния по концентрациям субстрата, биомассы и ферментативных продуктов. Множественность стационарных состояний не ординарное явление в химической и биологической кинетике. Системы с ингибированием субстратом в открытом режиме — один из классических примеров такого рода систем, в которых могут иметь место несколько стационарных состояний. Для такого рода систем наблюдается ряд весьма интересных особенностей в кинетическом поведении, например гистерезисные зависимости скорости реакции от скорости протока в реакторе.
Система может оказаться в одном из двух устойчивых стационарных состояний. Ответ на вопрос, в каком из двух таких состояний окажется система, достаточно сложен в теоретическом плане и зависит от предыстории системы. Только экспериментально можно ответить на этот вопрос. Как следует из уравнений (16.39), (16.40); (16.45), (16.46) и рис. 16.6, 16.7, увеличение скорости разбавления для системы, находящейся в состоянии 1, приводит к уменьшению концентрации субстрата и соответственно к росту концентрации биомассы или продукта реакции. В то же время увеличение скорости протока для системы, находящейся в состоянии 2, должно приводить к увеличению стационарного уровня субстрата и соответственно к уменьшению стационарного уровня биомассы или продукта. Это может служить экспериментальным критерием в выборе «ветви», на которой находится система.
Как идентифицировать эффект ингибирования субстратом в каждом конкретном случае, отличить данный механизм от механизма неосложненного роста? Качественным указанием на то, что в системе имеет место ингибирование субстратом, может служить гистерезисное поведение системы: при переходе от одной скорости разбавления к другой и обратно система может не вернуться к исходным значениям по концентрации компонентов процесса. Отклонения могут достигать больших значений.
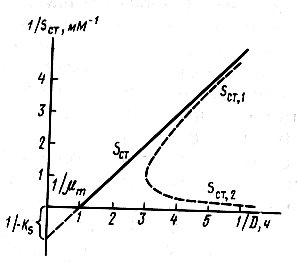
Количественные оценки зависимости стационарных концентраций компонентов от скорости разбавления для случая ингибирования избытком субстрата существенно отличаются от случая неосложненного роста (рис. 16.8). Как следует из рисунка, для неосложненного роста зависимость 1/Sст от 1/D представляет собой прямую, отсекающую на осях значения 1/μm и —1/KS. Данные для роста культуры с ингибированием в процессе реакции не должны линеаризоваться в обратных координатах. При этом если система функционирует, находясь на первой «ветви» стационарных концентраций (значения Sст2), то увеличение скорости разбавления должно приводить к уменьшению стационарной концентрации субстрата. Это кардинально отличает два обсуждаемых механизма роста.
|
Рис.16.8. Стационарные состояния в хемостате в случае неосложненного процесса (Sст) и в случае ингибирования избытком субстрата Sст1 и Sст2 в обратных координатах |
Определение
кинетических параметров роста культуры
из данных
по непрерывному культивированию
микроорганизма в
условиях ингибирования субстратом в
общем виде в силу множественности
стационарных состояний и иррациональности
основных
уравнений представляет определенные
сложности. В
качестве методического приема при
определении параметров можно
рекомендовать изучение стационарных
уровней системы в
условиях малых скоростей разбавления
в режиме D<<Dc.
В этих условиях
при использовании уравнений (16.39), (16.40)
и (16.45), (16.46) могут
быть найдены числовые значения параметров
![]() и
и
![]() .
.
Для полного анализа и определения всех параметров процесса уместным предствляется изучение кинетических кривых роста микроорганизмов при различных начальных концентрациях субстрата в режиме периодического культивирования.