

Лекция № 15. Периоды индукции на кинетических кривых роста микроорганизмов
Часто наблюдаемые периоды (лаг-фаза) индукции на кинетических кривых роста микроорганизмов могут иметь различную природу. Можно представить себе по крайней мере три механизма, реализация которых приводит к появлению на кинетической кривой роста популяции периода, в течение которого не происходит увеличения числа клеток или заметного образования продуктов реакции. Следует подчеркнуть, что период индукции трудно обнаружить из зависимости числа клеток или количества продуктов от времени при прямом наблюдении за этими параметрами, поскольку логарифмический рост культуры приводит к появлению кажущейся лаг-фазы. Период индукции τ определяется при анализе экспоненциальной фазы роста в полулогарифмических координатах из зависимости ln(N/N0) от τ как отрезок, отсекаемый на оси абсцисс при n(N/N0) = 0.
Период индукции может отражать следующие процессы.
1. Неспецифическую, неферментативную трансформацию исходного субстрата в продукт, приемлемый для дальнейшего метаболизма микроорганизмом:
![]() (15.1)
(15.1)
Вещество S' не используется микроорганизмом в силу его химической природы или физического состояния. Процесс перехода вещества S' в субстрат S характеризуется константой скорости k0 и может представлять собой химическое превращение вещества или его физическую трансформацию (например, переход из одной фазы в другую).
2. Адаптационные процессы в клетке, в первую очередь синтез необходимого фермента или ферментной системы:
![]() (15.2)
(15.2)
Состояние клеток N' характеризуется отсутствием необходимой ферментной системы и неспособностью клетки усваивать тот или иной субстрат и проводить акт деления. Образовавшаяся в результате адаптационного процесса клетка в состоянии N способна к автокаталитическому росту:
N→2N (15.3)
3. Присутствие в культуралъной среде ингибитора роста, блокирующего развитие популяции. С течением времени концентрация ингибитора I может падать в результате его расхода или вывода из системы:
![]() (15.4)
(15.4)
В качестве типичного примера данного механизма, вызывающего период индукции, можно привести влияние кислорода на рост анаэробных микроорганизмов. В присутствии заметных количеств кислорода рост популяции блокирован, однако за счет процессов восстановления кислорода или вытеснения его в газовую фазу системы «избавляется» от сильного ингибитора и начинается рост микробной популяции.
Три обсуждаемых механизма, приводящих к возникновению периода индукции, могут быть дифференцированы путем кинетического анализа и эксперимента.
15.1. Трансформация пресубстрата в субстрат
В том случае, если накопление субстрата в системе идет в соответствии со схемой (15.1), кинетику его роста описывает уравнение
![]() (15.5)
(15.5)
интегрирование которого дает
![]() (15.6)
(15.6)
где S0—начальная введенная в систему концентрация вещества S'. Если зависимость скорости роста микроорганизма от концентрации субстрата определяется классическим уравнением Моно, то дифференциальное уравнение роста будет иметь вид
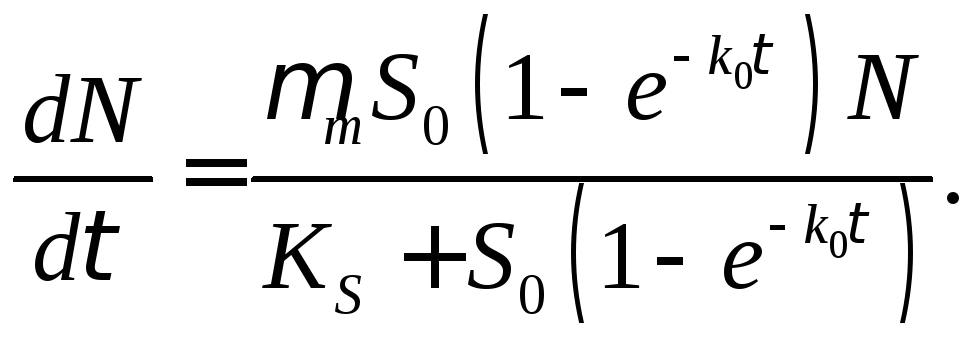 (15.7)
(15.7)
Это уравнение с разделяющимися переменными:
 (15.8)
(15.8)
Интегрирование этого уравнения дает
 (15.9)
(15.9)
Из уравнения (15.9) следует, что при τ=0 ln(N/N0) = 0 или N = N0, т. е. уравнение (15.9) справедливо при данных начальных условиях.
В процессе развития системы экспоненциальный член, входящий в логарифмическую часть уравнения (15.9), уменьшается и достигает уровня, когда справедливо неравенство
![]() (15.10)
(15.10)
При этом условии уравнение (15.9) трансформируется к виду
![]() (15.11)
(15.11)
где
![]() (15.12)
(15.12)
Это уравнение прямой, тангенс угла наклона которой представляет собой нечто иное, как удельную скорость роста микроорганизма:
![]()
В точке ln(N/N0) = 0 имеем
![]() (15.13)
(15.13)
или
![]() (15.14)
(15.14)
Таким образом, на кинетической кривой роста культуры в случае наличия в механизме процесса кинетической стадии трансформации исходного вещества в приемлемый метаболит должен наблюдаться период индукции. Протяженность периода индукции связана с начальной концентрацией предсубстрата соотношением
 (15.15)
(15.15)
Из уравнения (15.15) следует важная особенность кинетики процесса для механизма, включающего стадию трансформации предсубстрата: высокие концентрации предсубстрата могут полностью элиминировать период индукции.
Рассмотрим два частных случая.
1. Если начальная концентрация предсубстрата очень мала по сравнению с константой КS, S0 <<КS, то
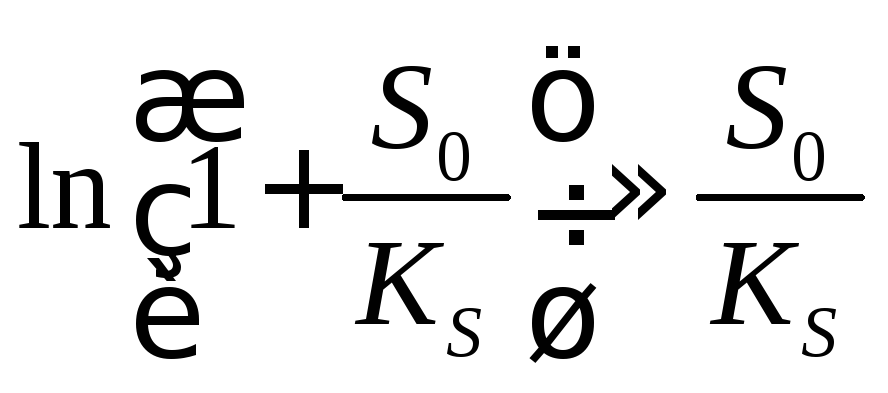 (15.16)
(15.16)
и
![]() (15.17)
(15.17)
Период индукции не зависит от концентрации субстрата и определяется только константой скорости трансформации предсубстрата в субстрат.
2. Если начальная концентрация предсубстрата существенно превышает значение константы KS , S0>>KS, то имеем
![]() (15.18)
(15.18)
Период индукции существенно уменьшается с увеличением начальной концентрации предсубстрата. В пределе при S0→∞ τs→0, т. е. при больших концентрациях субстрата период индукции отсутствует.