

Лекция № 13. Ингибирование роста популяции микроорганизмов продуктами ферментации
Один из наиболее часто встречающихся эффектов, приводящих к отклонению кинетики роста микроорганизмов от экспоненциальной кривой, связан с ингибированием их роста продуктами ферментативных превращений. Накопление какого-либо одного продукта превращения субстрата или их совокупности способно приводить к заметному уменьшению скорости роста культуры, переходу ее в стадию стационарного роста и даже к полной остановке процесса.
В качестве иллюстрации можно указать, что наиболее типичным случаем ингибирования роста микроорганизма продуктами жизнедеятельности является случай накопления в культуре органических кислот, снижающих значение рН среды культивирования. Как известно, скорость ферментативных процессов и, как следствие, скорость роста микроорганизмов рН-чувствительны. Если протоны, взаимодействуя с функционально важными компонентами клетки, блокируют метаболические процессы, феноменологически это означает, что они выступают в качестве ингибиторов роста.
Другим характерным примером могут быть процессы, протекающие с накоплением органических продуктов, в частности спиртов. Как правило, спиртовое брожение останавливается при накоплении в системе определенной концентрации продукта. Эффекты такого рода весьма распространены, в силу этого они требуют тщательного кинетического анализа.
Рассмотрим кинетику роста микробной популяции в приближении, учитывающем кинетически дискриминированные два процесса: стадию взаимодействия субстрата с клеткой и стадию деления клеток.
13.1. Ингибирование продуктом на стадии взаимодействия субстрата с клеткой
Если продукт реакции Р образуется на второй стадии, а процесс ингибирования заключается во взаимодействии продукта с формой N, то кинетическую схему процесса можно представить в виде
 (13.1)
(13.1)
Качественно ингибирование процесса продуктом заключается в том, что образуется его комплекс с активной формой N, при этом частица NР теряет способность к осуществлению последующего превращения в X и дальнейшему делению клетки. При развитии процесса с накоплением продукта все большая и большая доля N должна уводиться из реакционноспособного состояния, что должно тормозить процесс роста культуры.
В режиме постоянства концентрации субстрата (S=S0) переменными во времени компонентами системы являются частицы X, Р и NР. Эти переменные связаны между собой системой дифференциальных уравнений
![]() (13.2)
(13.2)
![]() (13.3)
(13.3)
![]() (13.4)
(13.4)
Сложение этих уравнений с учетом того, что общая биомасса М является суммой концентраций отдельных составляющих, т. е.
![]() (13.5)
(13.5)
приводит к соотношению
![]() (13.6)
(13.6)
Если
продукт Р
выступает в качестве быстрого и обратимого
ингибитора (это означает, что![]() ),
то процесс образования
комплекса активного центра какого-либо
фермента с
продуктом может быть охарактеризован
константой равновесия:
),
то процесс образования
комплекса активного центра какого-либо
фермента с
продуктом может быть охарактеризован
константой равновесия:
![]() (13.7)
(13.7)
В этом режиме динамика накопления продукта описывается уравнением
![]() (13.8)
(13.8)
где Yр — стехиометрический коэффициент, характеризующий количество продукта на единицу клеточной биомассы.
Из уравнений (13.6) и (13.8) следует весьма важное соотношение
![]() (13.9)
(13.9)
Это означает, что продукт реакции и биомасса связаны между собой простой линейной связью
Yр(Р-Р0) = (М-М0) или, если Р0<<Р, N0<<М, то YрР≈М. (13.10)
Для дальнейшего анализа воспользуемся условиями малости параметра при производной в уравнении (13.3). Если значение константы скорости процесса k2 достаточно велико, то в уравнении (13.3) можно пренебречь членом (1/k2)dХ/dτ по сравнению с X. Это приводит к следующему соотношению между переменными:
![]() (13.11)
(13.11)
С учетом (13.11) уравнение (13.5) можно записать в виде
 (13.12)
(13.12)
Если учесть, что переменные Р и М связаны линейно по уравнению (13.10), то будем иметь
 (13.13)
(13.13)
Подставляя это выражение в (13.6), получим дифференциальное уравнение, описывающее динамику изменения в системе общего количества клеток или общей биомассы:
 (13.14)
(13.14)
Уравнение (13.14) может быть записано в форме
![]() ,
(13.15)
,
(13.15)
где
 (13.16)
(13.16)
Видно, что по своему содержанию уравнение (13.16) соответствует случаю конкурентного ингибирования продуктом. Это уравнение с разделяющимися переменными
 (13.17)
(13.17)
которое может быть достаточно просто проинтегрировано:
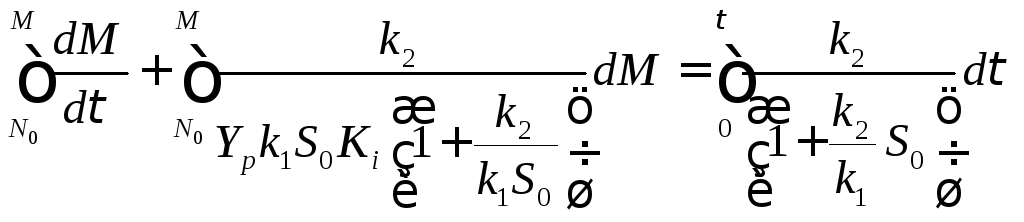 (13.18)
(13.18)
или
![]() (13.19)
(13.19)
Если процесс развивается достаточно глубоко, то в уравнении (13.19) можно без особой погрешности пренебречь N0 по сравнению с М и тогда оно примет вид
![]() (13.20)
(13.20)
Следует
обратить внимание на то, что множитель
перед τ
в
правой части уравнения (13.20) представляет
собой удельную скорость роста
микроорганизма в отсутствие процесса
ингибирования
продуктом![]() С учетом этого, уравнение (13.20)
можно записать так:
С учетом этого, уравнение (13.20)
можно записать так:
![]() (13.21)
(13.21)
Система будет функционировать в двух принципиально различающихся крайних режимах.
1. Если соотношения параметров уравнения (13.21) таковы, что в исследуемом интервале времени
![]() (13.22)
(13.22)
(например, исключительно велика константа ингибирования, (Кi-→∞), то кинетика роста популяции микроорганизма имеет обычный экспоненциальный вид
![]() (13.23)
(13.23)
и процесс ингибирования продуктом никоим образом не отражается на кинетике роста популяции. Эффекты ингибирования пренебрежимо малы.
2. Если соотношения параметров уравнения (13.21) таковы, что имеет место обратное неравенство
![]() (13.24)
(13.24)
(мало значение Кi или k1S0), то кинетика роста культуры не описывается экспоненциальной кривой и должно наблюдаться линейное увеличение биомассы:
![]() (13.25)
(13.25)
Для выявления необходимых соотношений между параметрами, регулирующих направление неравенств (13.22) и (13.24), запишем (13.22) в виде
 (13.26)
(13.26)
Поскольку сравнивать функции, указанные в правой и левой частях неравенства, достаточно
неудобно, сравним их производные в точке М = 0, так как неравенство производных в этой точке практически однозначно приводит к неравенству функции при произвольном М. Дифференцирование правой и левой частей неравенства (13.26) по переменной М в точке М = 0 приводит к выражению
![]() (13.27)
или
(13.27)
или
![]() (13.28)
(13.28)
Соотношение (13.28) может быть записано как
![]() (13.29)
(13.29)
С учетом того что в рамках используемой схемы k2 /k1 =КS, неравенство (11.29) будет иметь вид
![]() (13.30)
(13.30)
Из этих неравенств следует, что экспоненциальный или линейный рост микробиологической системы зависит от многих параметров: начальной концентрации клеток, удельной скорости роста, концентрации субстрата. Однако наиболее важным является значение Кi. На качественном уровне можно утверждать, что чем меньше Кi, тем больше значение коэффициента перед линейным членом уравнения (13.21) и тем сильнее проявляются эффекты ингибирования продуктом.
Введем безразмерную переменную M/N0. Уравнение (13.21) при соизмеримых M и N0 можно записать в виде
 (13.31)
(13.31)
или
 (13.32)
(13.32)
где φ=N0μ/(Kik1S0Yp). Это уравнение может быть представлено рядом линейных модификаций:
![]() (13.33)
(13.33)
 (13.34)
(13.34)
 (13.35)
(13.35)
Если механизм процесса сопряжен с ингибированием продуктом реакции, то экспериментально наблюдаемые зависимости должны линеаризоваться в координатах уравнений. Например как следует из уравнения (13.33), должна существовать линейная зависимость экспериментально определяемой величины (1/τ)·ln(M/N0) от (M/N0 – 1)/τ. В соответствии с уравнением (13.33) отрезок, отсекаемый этой линейной зависимостью на оси ординат, равен удельной скорости роста; тангенс угла зависимости дает параметр φ.
Величина φ является безразмерным параметром и характеризует эффекты ингибирования продуктом. Если экспериментально найденное значение φ близко к нулю, то можно утверждать, что в исследуемой системе отсутствуют эффекты ингибирования продуктом. Если значение φ в рамках статистической ошибки отлично от нуля, это говорит о заметном вкладе ингибирования продуктом в кинетику роста популяции. Значение константы ингибирования Ki можно определить из уравнения для φ, если известны параметры удельной скорости роста и значение N0/Yp.