

Лекция №12. Ингибирование роста популяции микроорганизмов избытком субстрата
Одной из характерных особенностей роста микроорганизмов являются часто наблюдаемые эффекты его ингибирования избытком субстрата. Удельная скорость роста культур микроорганизмов на начальном участке зависимости растет с увеличением концентрации субстрата, выходит на «насыщение», а затем при высоких концентрациях субстрата начинает уменьшаться. В силу этого активный рост популяции становится возможным лишь в узком диапазоне концентраций субстрата. Феномены такого рода требуют тщательного анализа и объяснения.
Простейший кинетический механизм, объясняющий эффект ингибирования автокаталитической реакции избытком субстрата, может быть представлен схемой
 (12.1)
(12.1)
Если в рамках химико-кинетического приближения записать систему уравнений для каждого из состояний N, X и ХS, концентрация которых изменяется в ходе процесса, то получим систему уравнений при условии, что концентрация субстрата постоянна и равна S0:
![]() (12.2)
(12.2)
![]() (12.3)
(12.3)
![]() (12.4)
(12.4)
Общее количество биомассы М в системе равно
M=N+X+XS (12.5)
или
![]() (12.6)
(12.6)
Подставив уравнения (12.2) — (12.4) в (12.6), получим
![]() (12.7)
(12.7)
Если процессы на обратимых стадиях схемы (12.1) протекают относительно быстро и эти реакции имеют равновесный характер, то можно выразить связи между переменными X, N и ХS с помощью уравнений для констант равновесия
![]() (12.8)
(12.8)
![]() (12.9)
(12.9)
где
![]() и
и
![]() —
равновесные константы диссоциации
комплексов.
—
равновесные константы диссоциации
комплексов.
В этом случае уравнение (12.5) может быть записано в виде
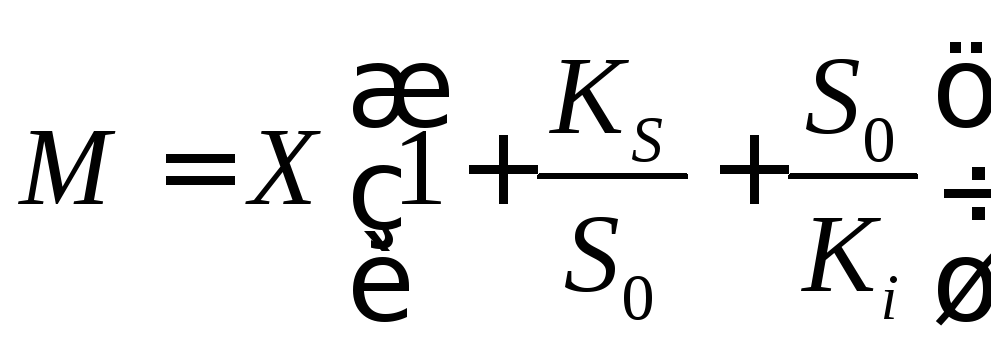 (12.10)
(12.10)
или
 (12.11)
(12.11)
Подстановка этого соотношения в уравнение (12.7) приводит к дифференциальному уравнению
 (12.12)
(12.12)
Решение его при использовании начального условия τ= 0, М = N0 имеет вид
![]() (12.13)
(12.13)
Общее число клеток экспоненциально увеличивается во времени, при этом удельная скорость роста должна быть представлена уравнением
 (12.14)
(12.14)
Из схемы (12.1) следует, что k=μm , тогда зависимость удельной скорости роста от концентрации субстрат при ингибировании его избытком запишется
 (12.15)
(12.15)
Рассмотрим кинетику процесса до полного исчерпания субстрата. Дифференциальное уравнение роста культуры в условиях ингибирования субстратом имеет вид
 (12.16)
(12.16)
где S — текущая концентрация субстрата; KS и Ki — эффективные значения субстратной константы и константы ингибирования. Это уравнение после разделения переменных можно записать в форме
![]() (12.17)
(12.17)
С
учетом балансового соотношения
![]() ,
а также
,
а также![]() ,
,![]() и приτ=0
N=N0
интеграл уравнения
(12.17) имеет вид
и приτ=0
N=N0
интеграл уравнения
(12.17) имеет вид
 (12.18)
(12.18)
Видно, что отличие этого уравнения от уравнения для процесса в отсутствие ингибирования субстратом заключается в дополнительном члене при множителе первого логарифмического слагаемого и в дополнительном отрицательном линейном члене. Из уравнения (12.18) следует, что при Кi→∞ (отсутствие или очень «плохое» ингибирование субстратом) уравнение (12.18) трансформируется в обычное интегральное уравнение роста культуры [см. (11.27)].
Уравнение (12.18) можно преобразовать к виду

(12.19)
Правая часть выражения (12.19) представляет собой удельную скорость роста культуры в экспоненциальной фазе. Действительно, когда величина N близка к N0(N-N0≈0), уравнение (12.19) трансформируется к
![]() (12.20)
(12.20)
где
 (12.21)
(12.21)
Удельная скорость роста культуры в этом случае является экстремальной функцией концентрации субстрата: при низких концентрациях субстрата скорость роста культуры растет с увеличением его концентрации, при высоких - падает. Для того чтобы выявить ингибирование процесса роста культуры избытком субстрата, в первую очередь необходимо исследовать его динамику в экспоненциальной фазе и показать соответствие экспериментальных данных уравнению (12.21).
Если разложить второй логарифмический член уравнения (12.19) в степенной ряд и использовать первый линейный член разложения, то уравнение (12.19) преобразуется к виду

(12.22)
Если
учесть, что![]() ,
то можно отрицательный член этого
уравнения представить в виде
,
то можно отрицательный член этого
уравнения представить в виде
 (12.23)
(12.23)
Таким образом, уравнение (12.19) можно записать так:
 (12.24)
(12.24)
где
 (12.25)
(12.25)
Следовательно, в этом уравнении удалось сгруппировать переменные таким образом, чтобы экспериментальные данные можно было представить в виде линейной анаморфозы, при этом параметры Ф и μ зависят от начальной концентрации субстрата, что позволяет идентифицировать случай развития процесса с ингибированием избытком субстрата.
Параметры μ и Ф могут быть найдены по известной процедуре линеаризации экспериментальных данных в координатах уравнения
 (12.26)
(12.26)
Как видно из уравнения (12.25), параметр Ф весьма сложно зависит от начальной концентрации субстрата. Можно выделить три области этой зависимости.
1.
![]()
При этом
![]() (12.27)
(12.27)
Параметр Ф не включает в себя константу ингибирования, и, следовательно, ингибирование избытком субстрата не обнаруживается.
2.
![]() Здесь
проявляется максимальная
независимость удельной скорости роста
от концентрации субстрата,
при этом Ф≈0, а микробная популяция
развивается строго
экспоненциально вплоть до глубины
конверсии 0,6—0,8.
Здесь
проявляется максимальная
независимость удельной скорости роста
от концентрации субстрата,
при этом Ф≈0, а микробная популяция
развивается строго
экспоненциально вплоть до глубины
конверсии 0,6—0,8.
3.
![]() В
этой области Ф имеет отрицательное
значение:
В
этой области Ф имеет отрицательное
значение:
![]() (12.28)
(12.28)
а скорость роста культуры увеличивается по мере израсходования субстрата; накопление продукта реакции и рост клеток опережают экспоненциальный рост.
В общем случае при анализе зависимости Ф от концентрации субстрата можно использовать трансформированное уравнение (12.25)
 (12.29)
(12.29)
Необходимо обратить внимание, что
 (12.30)
(12.30)
С учетом соотношения (12.30) уравнение (12.29) запишется как
![]() (12.31)
(12.31)
Уравнение (12.31) можно представить следующим образом:
![]() (12.32)
(12.32)
при
этом весь пул измерений при росте культур
микроорганизмов
в условиях ингибирования субстратом
должен линеаризоваться в координатах
зависимости
![]() от
от![]() .Из
этой зависимости определяются параметры
КS
и
Ki
.
.Из
этой зависимости определяются параметры
КS
и
Ki
.
При анализе экспериментальных данных, если есть основания считать, что процесс может ингибироваться избытком субстрата, можно рекомендовать следующую процедуру. Необходимо исследовать зависимость накопления клеток (или продуктов реакции) в экспоненциальной фазе при различных начальных концентрациях субстрата с определением ц известными и описанными ранее способами. Если зависимость обнаружит экстремальный характер, это будет однозначно свидетельствовать в пользу процесса с ингибированием избытком субстрата.
Однако информацию о том, что развитие микроорганизма связано с ингибированием избытком субстрата, можно получить и из анализа одной-единственной кинетической кривой роста. Если использовать линеаризацию экспериментальных данных в координатах уравнения (12.26) до степени конверсии 0,4—0,6, то можно определить значения μ и Ф. В том случае, если найденное Ф отрицательно, можно с уверенностью утверждать, что процесс протекает с ингибированием избытком субстрата.