

ХРЯЩЕВАЯ, КОСТНАЯ И МЫШЕЧНАЯ ТКАНИ
.pdfThis version of Total HTML Converter is unregistered.
филаменты, так называемые поперечные мостики. Положения их меняется при сокращении мышечного волокна.
Саркоплазматическая сетка и Т -система.
Каждая манжета состоит из трех компонентов:
1 ) терминальных цистерн (это плоские резервуары с краев манжеты);
2) саркотубул (трубочек, отходящих от терминальных цистерн и идут навстречу одни к другим );
3 ) центральной части, где саркотубулы образуют многочисленные анастомозы, напоминающие кружево. В целом описан элемент саркоплазматической сети имеет вид драной манжеты ( рваного рукава ). У млекопитающих терминальные цистерны проходят на границе А- и I- дисков саркомеров и поэтому в одном саркомере расположен один целый элемент ( манжета) на уровне диска А и половины двух соседних. Иначе говоря, элементы саркоплазматической сетки окружают А - диски, чередуются с элементами, окружающих И -диски. Элементы вокруг И - диска охватывают концевые участки соседских саркомеров.
Саркоплазматическая сетка
Между двумя соседними терминальными цистернами ретикулума расположена поперечная трубочка (Т - трубочка, или Т -
система). Т - трубочка - это система узких канальцев которые идут от плазмолеммы мышечного волокна (как ее вгинання) в
поперечном направлении примерно на равных расстояниях. Всередине волокна T - трубочки широко разветвляются. В мышцах млекопитающих ветви двух Т - трубочек оточуют каждый саркомер на границе между А- и I- дисками и контактируют, как уже было упомянуто, с двумя терминальными цистернами саркоплазматической сетки, образуя при этом так называемую триаду.

This version of Total HTML Converter is unregistered.
Последняя включает одну трубочку и две цистерны. Значение Т - системы заключается в том, что по ней нервный импульс плазмолеммы проникает в глубину мышечного волокна, включая все миофибриллы. Нервный импульс вызывает изменение проникливости мембран саркоплазматической сетки и выход вследствие этого ионов кальция в саркоплазму, где они необходимы для сокращения миофибрилл. Во время расслабления мышцы саркоплазматической сетки обеспечивает обратный транспорт ионов кальция от миофибрилл к своим полостям, используя для этого фермент АТФ - азу .
Молекулярные механизмы сокращения мышечного волокна.
Сучасные знания о механизме сокращения мышечного волокна базируются на представлении о филаментах двух типов,
которые сдвигаются одни относительно других. Эти представления является основой модели скользящих нитей, предложенной Г. Хаксли с сотрудниками на базе электронно - микроскопических исследований. Чтобы выяснить механизм взаимодействия актиновых и миозиновых филаментов, следует рассмотреть их молекулярное строение .
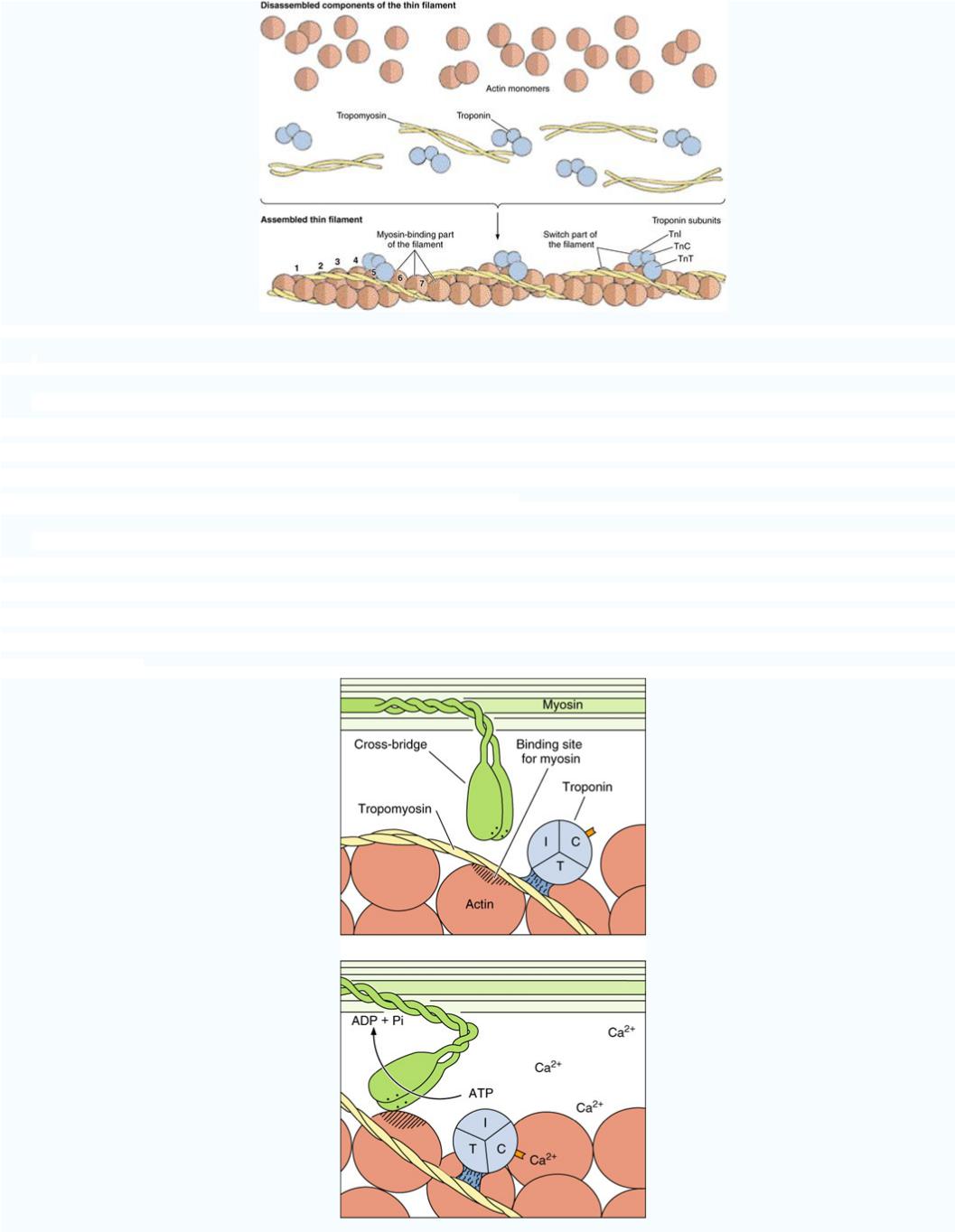
Тонкий филамент являет собой двойную спираль, построенную из двух цепочек глобулярных молекул актина (остов филамента). В продольных спиральных желобках с обеих сторон от актиновых цепочек лежат молекулы тропомиозина. К
молекулам тропомиозина на определенных расстояниях друг от друга присоединены молекулы тропонина. Тропомиозин вместе с тропонином играет основную роль в регуляции взаимодействия актина с миозином.
This version of Total HTML Converter is unregistered.
Толстые филаменты состоят из молекул миозина. Каждая молекула имеет двойную головку и длинный хвост и может сгибаться в двух местах так, что головка и проксимальная часть хвоста способны вращатся, как на шарнире. В толстом филаменте молекулы миозина лежат параллельно, образуя пучок. Половина их обращена головками к одному концу филамента, а
вторая - в другой. Молекулы миозина несколько сдвинуты друг относительно друга и их головки распологаются вдоль толстого филамента, исключая его серединную часть, где головок нет совсем .
Срединная часть толстого филамента построена только из хвостов миозиновых молекул. На электронных микрофотографиях головкам молекул миозина отвечают вышеупомянутые поперечные мостики, которые во время сокращения мышечного волокна образуют многочисленные соединения между толстыми и тонкими филаментами. Головки миозина расположенны по спирали,
образуя шесть продольных рядов. Каждый ряд головок лежит точно против одного из шести тонких филаментов, которые окружают один толстый филамент. Во время сокращения, головки миозина присоединяются к молекулам актина в соседнем тонком филаменте .
Комплексы тропонина и тропомиозина действуют как своеобразное молекулярное «замыкальное устройство», которое во время расслабления мышечного волокна не дает молекулам актина взаимодействовать с миозиновыми головками толстых филаментов. «Открывают» актин ионы кальция, которые освобождаются из полостей саркоплазматической сети при распространении импульса по Т - трубочках. После остановки стимуляции ионы кальция быстро транспортируются от миофибрилл к саркоплазматической сетки. Тогда актин снова замыкается и сокращения прекращаются. Механизм, с помощью которого ионы кальция «открывают» актин, связанный с их присоединением к тропонину: молекулы тропомиозина при этом

This version of Total HTML Converter is unregistered.
сдвигаются и открывают участки актина, способные взаимодействовать с головками миозина .
Энергию, необходимую для сокращения мышц, дает АТФ. Головки миозина способны связывать молекулы АТФ и имеют АТФ - азную активность (способны расщеплять АТФ). Энергия, которая высвобождается при этом используется на изгиб молекул миозина в «шарнирных» участках, их присоединение к актиновым филаментам и продвижение последних вдоль миозиновых. Комплекс актина с миозином и АТФ не стабилен и быстро розпадается на актин и миозин - АТФ. Очевидно,
поперечные мостики отделяются в тот момент, когда головки миозина связываетют молекулы АТФ. Согласно рассчетам этот цикл повторяется с огромной скоростью - 50-100 раз в секунду. Интересным есть тот факт, что после смерти, вследствие прекращения синтеза АТФ, в мышцах не остается молекул, которые вызвали б отделение миозина от актина и актиномиозиновий комплекс стабилизируется на несколько часов. Филаменты фиксируются в соединеном положении. Это состояние имеет название трупного одубивания и сохраняется к появлению аутолитических изменений, после чего мышцы становятся способными к пассивному расслаблению.
Красные и белые мышечные волокна.
В саркоплазме содержится растворимый пигментный белок миоглобин . По своему химическому строению этот белок очень близок к гемоглобину крови и также способен связывать кислород и отдавать его при необходимости. Миоглобин окрашивает мышечные волокна в красный цвет. Зависимо от содержания саркоплазмы (а, следовательно, и миоглобина), толщины и ферментного состава мышечные волокна подразделяют на красные, белые и промежуточные . Мышцы человека в основном содержат все три типа волокон, но их соотношения зависит от функции того или иного мышца. Красные волокна имеют незначительную толщину, большое количество миоглобина в саркоплазме, многочисленные митохондрии, богатые цитохромы.
Белые волокна толще, они содержат меньше миоглобина и митохондрий. Волокна третьего типа занимают промежуточное положение по этим показателям. Мышцы, в которых преобладают красные волокна, способны к более длительной активности,
чем мышцы, состоящие преимущественно из белых волокон. Потому что их саркоплазма хорошо приспособлена к обеспечению своих энергетических потребностей. Белые волокна способны сокращатся быстрее, чем красные, но они сравнительно быстро устают, так как не могут долго получать достаточное количество энергии.
Функциональные особенности поперечно – исчерчаемой мышечной ткани.
Споперечно - исчерчаемой мышечной ткани построены мышцы скелета человека, сокращение которых зависит от сознания,
вотличие от непроизвольного сокращения гладких мышц. Поперечно - исчерчаемым мышцам присущ так называемый тетанических тип сокращения, для которого характерны такие признаки: сокращение сильные, быстрые (сокращение мышечных
волокон в 10-25 раз быстрее, чем гладких мышечных клеток), не продолжительны. Исчерчаемые мязы быстрее устают и не могут находиться в состоянии сокращения так долго, как гладкие.
Строение мышцы как органа.
Отдельные поперечно - исчерчаемые мышечные волокна сочетаются соединительной тканью в орган, имеющий название мышцы. Тонкие слои рыхлой соединительной ткани между мышечными волокнами называют эндомизием. Ретикулярные и коллагеновые волокна эндомизия переплетаються с волокнами сарколеммы. На конце каждого мышечного волокна плазмолемма образует узкикие глубокие вгинання, в которые проникают коллагеновые и ретикулярные волокна.

This version of Total HTML Converter is unregistered.
Последние пронизывают базальную мембрану и образуют петлю, которая фиксируется к плазмолемме именно в том месте,
где с ней контактируют актиновые нити саркомеров. После выхода из базальной мембраны ретику лярные волокна
переплетаются с коллагеновыми, а последние переходят в сухожилия. Каждое мязовое волокно имеет самостоятельную
иннервацию и окружено сеткой гемокапилляров. Комплекс волокна с окружающими его элементами рыхлой соединительной
ткани является структурной и функциональной единицей скелетной мышцы и называется мионом.
Сердечная мышечная ткань
Гистогенез сердечной мышечной ткани. Источники развития сердечной мышечной ткани находятся в прекардиальной мезодерме. В гистогенезе возникают парные складчатые утолщения висцерального листка спланхнотома — миоэпикардиальные пластинки, содержащие стволовые клетки сердечной мышечной ткани. Последние путем дивергентной дифференцировки дают начало следующим клеточным дифферонам: рабочим, ритмзадающим (пейсмекерным), проводящим и секреторным кардиомиоцитам. Исходные клетки сердечной мышечной ткани — кардиомиобласты характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндоплазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролиферативным потенциалом. После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез. В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постнатальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов. Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофибриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов "конец в конец", "конец в бок" формируют клеточные комплексы — сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру.
Строение сердечной мышечной ткани. Структурно-функциональной единицей волокон есть кардиомиоциты — это
клетки, имеющие вытянутую прямоугольную форму. Длина рабочих кардиомиоцитов составляет 50-120 мкм, а ширина — 15-20
мкм. Одно-два ядра располагаются в центре клетки.
Сердечная мышечная ткань
Сердечная мышечная ткань
This version of Total HTML Converter is unregistered.
Периферическую часть цитоплазмы кардиомиоцитов занимают поперечноисчерченные миофибриллы, аналогичные таковым в симпластах скелетномышечного волокна. Однако каналы саркоплазматической сети и Т-системы менее отчетливо выражены. Кардиомиоциты отличаются большим количеством митохондрий, расположенных тесными рядами между миофибриллами. Снаружи миоциты покрыты сарколеммой, в составе которой выделяются плазмолемма и базальная мембрана.
Характерной особенностью ткани является наличие вставочных дисков на границе между контактирующими кардиомиоцитами.
Вставочные диски пересекают волокно в виде волнистой или ступенчатой линии и включают межклеточные контакты от простых, по типу десмо-сом и до щелевых (нексусов).
Сердечная мышечная ткань
http://uk.wikipedia.org/wiki/%D0%9C%D1%96%D0%BD%D0%B5%D1%80%D0%B0%D0%BB%D0%B8
Источники информации
1. Улумбеков Э.Ф., Чельшева Ю.А. Гистология, эмбриология. Цитология / Э.Ф. Улумбеков, Ю.А. Чельшева – М. : ГЕО ТАР. –
Медиа, 2007. – С. 145 - 192
2.Данилов Р. К. Гистология. Эмбриология. Цитология. : [учебник для студентов медицинских вузов] / Р. К. Данилов – М. : ООО
«Медицинское информационное агенство», 2006. – С. 152 – 177.
3.Гистология, цитология и эмбриология / [Афанасьев Ю. И., Юрина Н. А., Котовский Е. Ф. и др.] ; под ред. Ю. И. Афанасьева,
Н. А. Юриной. – [5-е изд., перераб. и доп.]. – М. : Медицина. – 2001. – С. 224 - 268
4.Кузнецов С. Л. Атлас по гистологии, цитологии и эмбриологии / Кузнецов С. Л., Н. Н. Мушкамбаров, В. Л. Горячкина. – М. :
Медицинское информационное агенство, 2002. – С. С. 81 - 98.
http://uk.wikipedia.org/wiki/%D0%97%D0%BE%D0%B1%D1%80%D0%B0%D0%B6%D0%B5%D0%BD%D0%BD%D1%8F:Bone-inside.jpg
http://uk.wikipedia.org/wiki/%D0%97%D1%83%D0%B1
Автор |
доц. Шутурма О.Я. http://uk.wikipedia.org/wiki/%D0%97%D1%83%D0%B1 |
http://uk.wikipedia.org/w/index.php? title=%D0%93%D0%B0%D0%B9%D0%BC%D0%BE%D1%80%D0%BE%D0%B2%D0%B0_%D0%BF%D0%B5%D1%87%D0%B5%D1%80%D0%B0&action=edit
http://uk.wikipedia.org/w/index.php? title=%D0%93%D0%B0%D0%B9%D0%BC%D0%BE%D1%80%D0%BE%D0%B2%D0%B0_%D0%BF%D0%B5%D1%87%D0%B5%D1%80%D0%B0&action=edit
http://uk.wikipedia.org/w/index.php? title=%D0%93%D0%B0%D0%B9%D0%BC%D0%BE%D1%80%D0%BE%D0%B2%D0%B0_%D0%BF%D0%B5%D1%87%D0%B5%D1%80%D0%B0&action=edit