

ХРЯЩЕВАЯ, КОСТНАЯ И МЫШЕЧНАЯ ТКАНИ
.pdfThis version of Total HTML Converter is unregistered.
Одновременно с дифференцированием клеток в остеоциты образуется межклеточное вещество и коллагеновые волокна. Расположенное между клетками и волокнами основное вещество уплотняется, формируются костные балки (перекладины). Клетки поверхности образуемой кости превращаются в остеобласты. Большинство костей скелета человека (за исключением покровных костей черепа) построены из пластинчатой костной ткани, то есть имеют костные пластинки толщиной от 4 до 15 мкм, которые состоят из остеоцитов и тонковолокнистого межклеточного вещества. Соединительнотканные волокна в толще каждой пластинки лежат параллельно и ориентированы в определенном направлении. В зависимости от расположения костных пластинок различают компактные и губчатые кости. В компактной кости пластинки располагаются в определенном порядке, образовывая сложные системы остеоны.
http://uk.wikipedia.org/wiki/%D0%9C%D1%96%D0%BD%D0%B5%D1%80%D0%B0%D0%BB%D0%B8
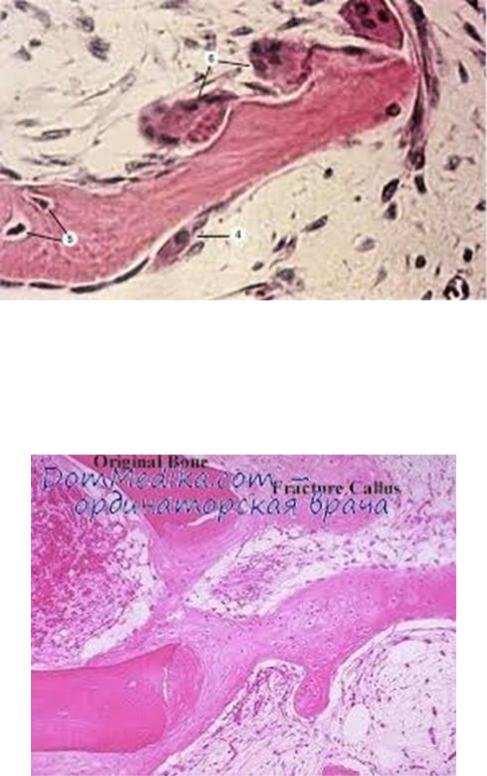
Гистогенез костной ткани.
Существует два способа развития костной ткани — непосредственно из мезенхимы (перепончатый остеогенез) и на месте хрящевого зачатка (хрящевой остеогенез). Первый способ характерен для первых недель эмбриогенеза, второй — для более поздних этапов эмбрионального развития и постнатального онтогенеза.
Из соединительной ткани развиваются кости свода и боковых отделов черепа, нижняя челюсть и, по мнению некоторых,
ключица (а у низших позвоночных и некоторые другие) — это так называемые покровные или облегающие кости. Они
развиваются прямо из соединительной ткани; волокна её несколько сгущаются, между ними появляются костные клетки и в
промежутках между последними отлагаются известковые соли; образуются сначала островки костной ткани, которые затем
сливаются между собой. Большинство костей скелета развивается из хрящевой основы, имеющей такую же форму, как будущая
кость. Хрящевая ткань подвергается процессу разрушения, всасывания и вместо неё образуется, при деятельном участии особого
слоя образовательных клеток (остеобластов), костная ткань; процесс этот может идти как с поверхности хряща, от одевающей
его оболочки, перихондрия, превращающегося затем в надкостницу, так и внутри его. Обыкновенно развитие костной ткани
начинается в нескольких точках, в трубчатых костях отдельными точками окостенения обладают эпифизы и диафиз.
This version of Total HTML Converter is unregistered.
При характеристике процессов развития костной ткани непосредственно из мезенхимы определяют следующие этапы. Первый этап — формирование в составе мезенхимы так называемого остеогенного зачатка. При этом проходит локальное размножение мезенхимных клеток с врастанием в скелетогенный островок кровеносных сосудов. Остеоидный, то есть второй этап, характеризуется выделением остеогенными клетками в межклеточное пространство коллагена (формированием осеиновых волокон) и высокомолекулярных биополимеров (гликопротеинов, протеoгликанов, липидов) осеомукоида. Третий этап — собственно образование грубоволокнистой кости — заключается в известковании межклеточного вещества (откладывании солей кальция). Для реализации этого процесса необходимо присутствие в межклеточном веществе продуцируемой остеобластами щелочной фосфатазы и белка остеонектина. Последний, связывая коллаген из гидроксиапатитом, определяет место роста кристаллов фосфата кальция и их прикрепление к органичному матриксу кости. Четвертый этап связан из резорбцией ткани заключается в постоянном изменении направления действия вектора силы на кость, в результате чего возникает, так называемый пьезоэлектрический эффект (устанавливается разница потенциалов на ввогнутой и выпуклых поверхностях костных пластинок). При чем отмечено, что концентрация остеобластов и процессы апозиционного новообразования кости связаны с отрицательными зарядами, а концентрация остеокластов и процессы резорбции — с позитивными зарядами на поверхности костной ткани.
Стадии перепончатого остеогенеза:
1.формирование остеогенного островка – костной бластемы;
2.остеоидная – формирование осеиновых волокон и осеомукоида;
3.образование грубоволокнистой костной ткани;
4.ЗАМЕНА ГРУБОВОЛОКНИСТОЙ НА ПЛАСТИНЧАТУЮ КОСТНУЮ ТКАНЬ.
СТАДИИ ХРЯЩЕВОГО ОСТЕОГЕНЕЗА:
1.образование хряща - на месте будущей кости образуется гиалиновый хрящ
2.перихондральное окостенение
o проходит только в области диафиза
o в области диафиза надхрящница превращается в надкостницу, в которой появляются остеогенные клетки - остеобласты
o за счет остеогенных клеток надкостницы на поверхности хряща начинается образование кости в виде общих пластинок, имеющих циркулярный ход, наподобие годовых колец дерева (см. пластинчатую кость)

This version of Total HTML Converter is unregistered.
3. эндохондральное окостенение
o происходит как в области диафиза, так и в области эпифиза; окостенение эпифиза осуществляется только путем эндохондрального окостенения
o внутрь хряща врастают кровеносные сосуды, в адвентиции которых имеются остеогенные клетки - остеобласты, за счет которых вокруг сосудов происходит образование кости в виде остеонов
oодновременно с образованием кости происходит разрушение хряща
4.перестройка и рост кости - старые участки кости постепенно разрушаются и на их месте образуются новые; за счет надкостницы образуются общие костные пластинки, за счет остеогенных клеток, находящихся в адвентиции сосудов кости, образуются остеоны
МЕТАЭПИФИЗАРНАЯ ПЛАСТИНКАИМЕЕТ4 ЗОНЫ;
1.НЕИЗМЕНЕННОГОХРЯЩА:
2.СТОЛБЧАТОГО ХРЯЩА;
3.ПУЗЫРЧАТОГО ХРЯЩА;
4.РЕЗОРБЦИИХРЯЩАИНЕЗРЕЛОЙКОСТНОЙТКАНИ.
РОСТ КОСТИ В ДЛИНУ ПРОИСХОДИТ ГЛАВНЫМ ОБРАЗОМ В ЧАСТЯХ ЕЩЕ НЕ ОКОСТЕНЕЛЫХ (В ТРУБЧАТЫХ КОСТЯХ МЕЖДУ ЭПИФИЗАМИ И ДИАФИЗОМ), НО ОТЧАСТИ И ПУТЕМ ОТЛОЖЕНИЯ НОВЫХ УЧАСТКОВ ТКАНИ МЕЖДУ СУЩЕСТВУЮЩИМИ, УТОЛЩЕНИЕ КОСТИ ПРОИСХОДИТ ПУТЕМ ОТЛОЖЕНИЯ НА ПОВЕРХНОСТИ КОСТИ НОВЫХ СЛОЕВ («АППОЗИЦИЯ») БЛАГОДАРЯ ДЕЯТЕЛЬНОСТИ ОСТЕОБЛАСТОВ НАДКОСТНИЦЫ. ПОСЛЕДНЯЯ ВЛАДЕЕТ В ВЫСОКОЙ СТЕПЕНИ СПОСОБНОСТЬЮ ВОСПРОИЗВОДИТЬ РАЗРУШЕННЫЕ И ИЗЪЯТЫЕ ЧАСТИ КОСТИ. ДЕЯТЕЛЬНОСТЬЮ ЕЕ ОБУСЛОВЛИВАЕТСЯ И СРАСТАНИЕ ПЕРЕЛОМОВ. ПАРАЛЛЕЛЬНО С РОСТОМ КИСТИ ИДЕТ РАЗРУШЕНИЕ, ВСАСЫВАНИЕ («РЕЗОРБЦИЯ») НЕКОТОРЫХ УЧАСТКОВ КОСТНОЙ ТКАНИ,
ПРИЧЕМ ДЕЯТЕЛЬНУЮ РОЛЬ ИГРАЮТ ТАК НАЗЫВАЕМЫЕ ОСТЕОКЛАСТЫ («КЛЕТКИ, КОТОРЫЕ РАЗРУШАЮТ КИСТЬ»), МНОГОЯДЕРНЫЕ ЭЛЕМЕНТЫ,
КОТОРЫЕ НАБЛЮДАЮТСЯ НАСТЕНКАХ МОЗГОВЫХ ПОЛОСТЕЙ, ВНАДКОСТНИЦЕ ИСТЕНКАХ БОЛЬШИХ ПОЛОСТЕЙ В КОСТИ (НАПР. ГАЙМОРОВАПАЗУХА И ТОМУ ПОДОБНОЕ
Изучение химического состава костной ткани сопряжено со значительными трудностями, поскольку для выделения органического матрикса требуется провести деминерализацию кости. Кроме того, содержание и состав органического матрикса подвержены значительным изменениям в зависимости от степени минерализации костной ткани.
Известно, что при продолжительной обработке кости в разведенных растворах кислот ее минеральные компоненты растворяются и остается гибкий мягкий органический остаток (органический матрикс), сохраняющий форму интактной кости.
Межклеточный органический матрикс компактной кости составляет около 20%, неорганические вещества – 70% и вода – 10%. В
губчатой кости преобладают органические компоненты, которые составляют более 50%, на долю неорганических соединений приходится 33–40%. Количество воды сохраняется в в тех же пределах, что и в компактной кости.
По данным А. Уайта и соавт., неорганические компоненты составляют около 1/4 объема кости; остальную часть занимает органический матрикс. Вследствие различий в относительной удельной массе органических и неорганических компонентов на долю нерастворимых минералов приходится половина массы кости.
Неорганический состав костной ткани. Более 100 лет назад было высказано предположение, что кристаллы костной ткани имеют структуру апатита. В дальнейшем это в значительной мере подтвердилось. Действительно, кристаллы кости
относятся к |
гидроксилапатитам, имеют форму пластин или |
палочек и следующий химический состав – |
Са10(РО4)6(ОН)2. |
Кристаллы гидроксилапатита составляют лишь часть |
минеральной фазы костной ткани, другая часть |
представлена аморфным фосфатом кальция Са3(РО4)2. Содержание аморфного фосфата кальция подвержено значительным колебаниям в зависимости от возраста. Аморфный фосфат кальция преобладает в раннем возрасте, в зрелой кости преобладающим становится кристаллический гидроксилапатит. Обычно аморфный фосфат кальция рассматривают как лабильный резерв ионов Са2+ и фосфата.
В организме взрослого человека содержится более 1 кг кальция, который почти целиком находится в костях и зубах,

This version of Total HTML Converter is unregistered.
образуя вместе с фосфатом нерастворимый гидроксилапатит. Большая часть кальция в костях постоянно обновляется.
Ежедневно кости скелета теряют и вновь восстанавливают примерно 700–800 мг кальция.
В состав минеральной фазы кости входит значительное количество ионов, которые обычно не содержатся в чистом
гидроксилапатите, например ионы натрия, магния, калия, хлора и др. Высказано предположение, что в кристаллической решетке гидроксилапатита ионы Са2+ могут замещаться другими двухвалентными катионами, тогда как анионы, отличные
от фосфата игидроксила, либо адсорбируются на поверхности кристаллов, либо растворяются в гидратной оболочке
кристаллической решетки.
ОСТЕОПОРОЗhttp://uk.wikipedia.org/wiki/%D0%9C%D1%96%D0%BD%D0%B5%D1%80%D0%B0%D0%BB%D0%B8
Вековые изменения костной ткани заключаются в постепенной потере неорганического матрикса кости после достижения двадцатилетнего возраста. Характерной особенностью является то, что у мужчин потеря минеральных компонентов кости является постоянным показателем на протяжении всей жизни и составляет около 0,4 % к массе ежегодно. У женщин с наступлением менопаузы, очевидно, в результате дефицита эстрогенов в организме, процессы деминерализации нарастают, достигая уровня 1–1,5 % ежегодно.
Количество (а точнее соотношение) органических и минеральных веществ в кости изменяется с возрастом. У детей значительно преобладают органические вещества, у взрослых минеральные. У взрослого человека часть минеральных составляющих (преимущественно, фосфорнокислая и углекислая известь, а также, хлористого кальция и другие) составляет около 60—70 % массы кости, а органичные вещества 40 %. Кости имеют большую прочность и сильное сопротивление разрыву, чрезвычайно долго противостоят разрушению. При сильном нагревании кисть теряет органические вещества, но хранит свою форму и строение; поддавая кисть действию кислоты (напр. серной), можно растворить минеральные вещества и получить гибкий хрящеватый скелет кости.
Мышечная ткань (rextus muscularis) построена из элементов, способных к сокращению, которые выполняют всю
совокупность двигательных процессов в организме (крово- и лимфообращение, передвижение пищи в пищеварительном тракте,
воздуха в дыхательных путях, работа сердца), а также перемещение организма или его частей в пространстве. Элементы
мышечных тканей содержат специальные органеллы - миофибриллы. В их основе лежат актиновые и миозиновые
миофиламенты, которые своим взаимодействием обеспечивают процесс сокращенияи, таким образом, осуществляят функцию
движения.
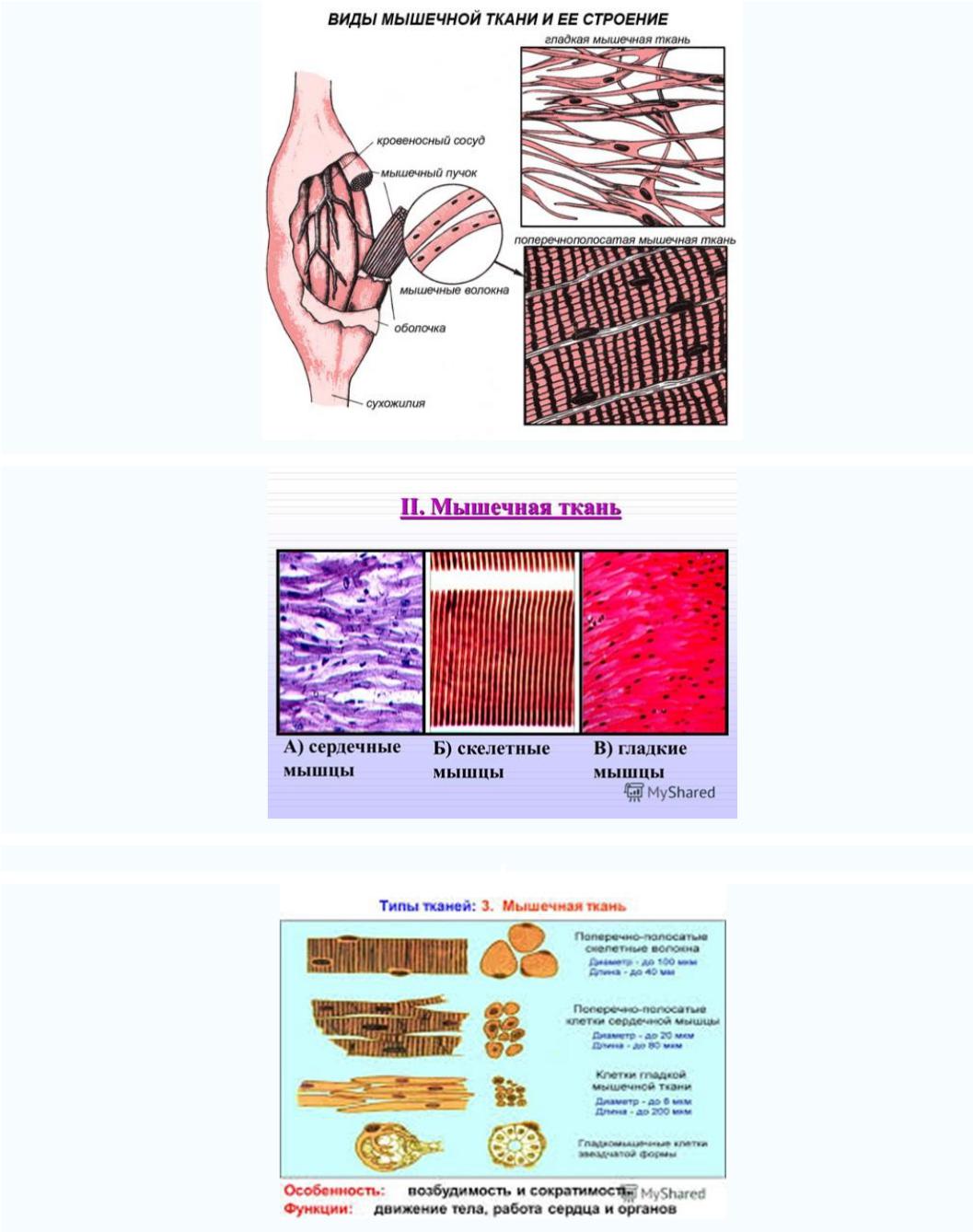
Существуют две классификации мышечных тканей - морфофункциональная и генетическая. Согласно морфофункциональной
классификацции мышечные ткани разделяют на две группы: гладкую и поперечно - исчерчаемую, которая, в свою очередь,
делится на скелетную и сердечню.
This version of Total HTML Converter is unregistered.
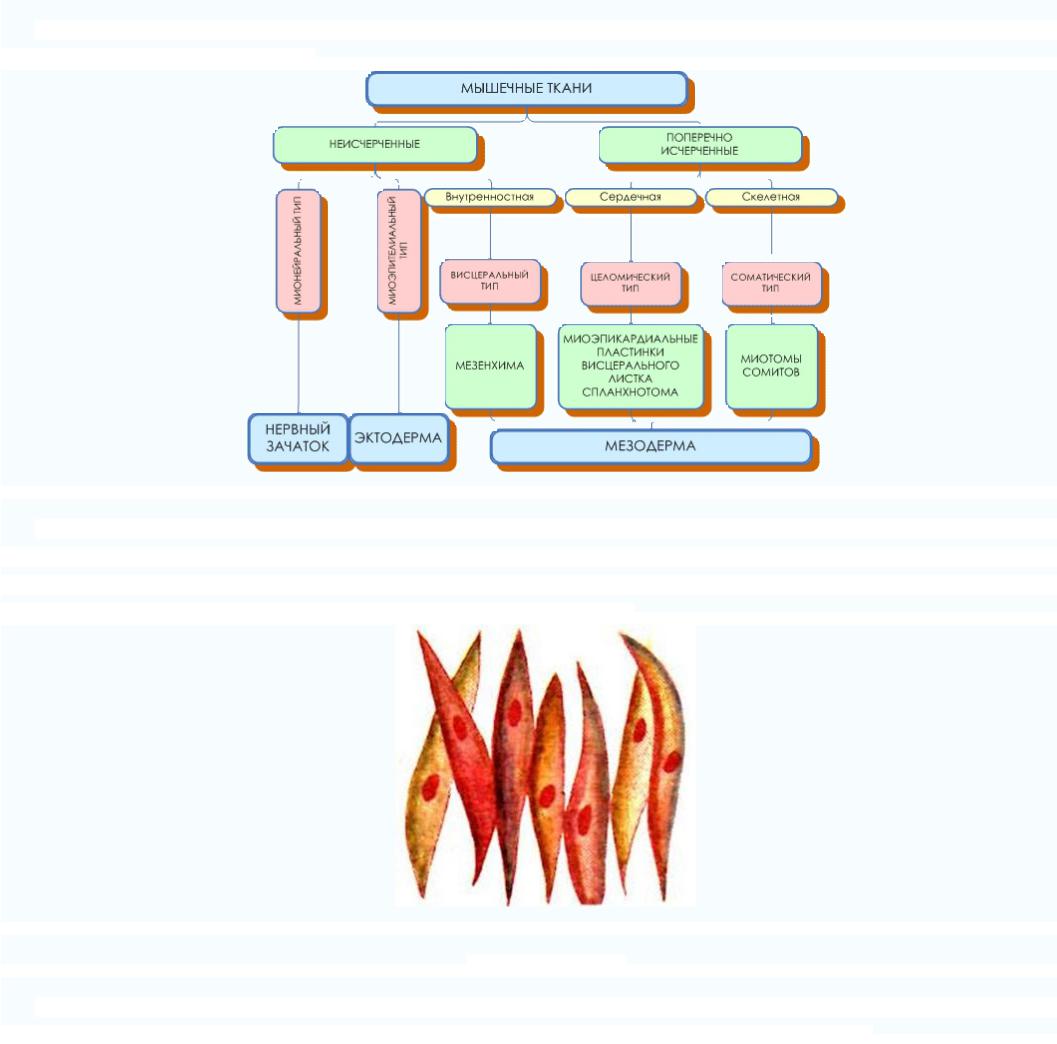
Согласно генетической классификации, которая была предложенна Н. Г. Хлопиным, мышечные ткани розделяются по их
происхождению на пять гистогенетических типов:
1)соматический тип происходит из миотомов мезодермы. Это скелетна мышечная ткань;
2)целомический тип происходит с вентральной мезодермы. Это сердечная мышечная ткань;
3)висцеральный тип происходит из мезенхимы. Это гладкая мышечная ткань внутренних органов;
4)невральный тип происходит из нервной трубки. К этому типу относятся гладкие миоциты мышц радужной оболочки
глаза;
This version of Total HTML Converter is unregistered.
5) эпидермальный тип происходит из кожной эктодермы, включает миоэпителиальные корзинообразное клетки потовых,
молочных, слюнных и слезных желез).
Гладкая мышечная ткань ( textus muscularis non striatus ) входит в состав стенок полостных внутренних органов, а также содержится в капсулах селезенки и лимфатических узлов, в коже. Происходит гладкая мышечная ткань из мезенхимы, то есть имеет общее происхождение с тканями внутренней среды. В группе мышечных тканей эта ткань рассматривается только с точки зрения строения и функции. Структурной единицей является гладкий миоцит.
Гладкие миоциты
Это веретенообразная клетка длинной от 20 до 100 мкм (в матке во время беременности она может достигать 500 мкм),
диаметром от 2 до 20 мкм. В матке, эндокарде, аорте, мочевом пузыре встречаются миоциты с отрастками.
Ядра миоцитов паличкообразной формы и лежат в центральной широкой части клеток. Оно содержит небольшое количество гетерохроматина, хорошо заметны ядрышка. Когда миоцит сокращается, ядро изгибается. Цитоплазма окрашивается оксифильно с базофильным оттенком. Органеллы общего назначения, среди которых много митохондрий,
содержатся у полюсов ядра. Комплекс Гольджи и эндоплазматическая сеть (особенно гранулярная ) развиты слабо. Цитоплазма содержит также включения - жировые, углеводные и пигментные. Цитоплазма имеет многочисленные вгинання - пиноцитозные пузырьки и кавеолы. С их помощью в цитоплазму поступают, в частности ионы кальция.

This version of Total HTML Converter is unregistered.
Гладкая мышечная ткань
Гладкая мышечная ткань
Миоциты не имеют поперечной исчерченности. Под электронным микроскопом в их цитоплазме выявляются тонкие
актиновые миофиламенты и толстые миозиновые, расположенные преимущественно продольно, но не так как в поперечно -
исчерчаемых мышцах и, очевидно, они не образовывают миофибрилл. Актиновых филаментов содержится больше. Они, кроме
продольного направления, идут под углом к оси клетки, образовывая объемную сетку. Фиксируются актиновые нити к
цитолеммы или друг к другу с помощью электронноплотные телец, которые состоят из белка альфа - актинина. Благодаря межмолекулярным взаимодействием с миозином актиновые нити передвигаются навстречу друг другу, тяга передается на цитолему и клетка сокращается. В механизме сокращения гладких миоцитов большую роль играет процесс фосфорилирования миозна, который зависит от концентрации ионов кальция. В свою очередь, регуляция концентрации этих ионов происходит с помощью специального белка, связывающего кальций - кальмодулина. Кальмодулин в комплексе с кальцием активирует фермент, что фосфорилирует миозин . В фосфорилированном состоянии миозин способен к взаимодействию с актином.
Оболочка каждого миоцита окутана тонкой базальной мембраной, к которой прикрепляются коллагеновые фибриллы. В
базальной мембране имеются отверстия , в области которых мышечные клетки контактируют друг с другом с помощью щелевых контактов (нексусов). Вокруг мышечных клеток ретикулярные, эластические и тонкие коллагеновые волокна образуют сетку -
эндомизий, который обьеденяет соседние миоциты. Мышечные группы по 10-12 мышечных клеток, в свою очередь,
объединяются в мышечные пласты, между которыми лежит рыхлая соединительная ткань с сосудами и нервами.

This version of Total HTML Converter is unregistered.
Сокращается гладкая мышечная ткань ритмично, медленно, но способна долго находиться в состояние сокращения, не уставая при этом. Медленное сокращение ее обусловлено медленным циклом взаимодействия миозина с акты ном. Гладкая мускулатура способна на большие силы сокращений (например, мышечная оболочка беременной матки при родах). Тип сокращение, свойственный гладким мышцам, называется тонический. Сокращение висцеральной мускулатуры является непроизвольным, т.е. не поддается контролю сознания.
Поперечно – исчерчаемая мышечная ткань.
Скелетная мышечная ткань занимает 42% массы тела взрослого человека, причем подавляющее большинство мышц образована скелетной мышечной тканью. Источником развития этой ткани являются клетки миотомов дорзальной мезодермы.
Эти клетки диференцирирюются в двух направлениях. Одни способны сливатся и строить симпластические структуры -
мышечные трубочки, которые далее формирую дефинитивное образования - миосимпласты. Вторая линия дает при развитии -
миосателитоциты.
Единицей строения скелетной мышечной ткани является мышечное волокно, образованное миосимпластом и миосателитоцитамы. Мышечное волокно имеет форму цилиндра, концы которого могут быть округлены, скошенные или зазубренные. Диаметр волокна 9-150 мкм (9 мкм новорожденного ребенка, 40-50 мкм у взрослого, 150 мкм спортсмена). Длина мышечного волокна часто совпадает с длиной мышцы и может быть различна в зависимости от размеров мышцы. Например, в
портняжной мышце человека она может достигать 12-13 см. Волокно окружено сарколеммой
Мышечные волокна

This version of Total HTML Converter is unregistered.
Мышечные волокна
Сарколемма состоит из внешней базальной мембраны. Внутренним слоем сарколеммы является плазмолемма миосимпласта.
Она участвует в проведении импульсов. Между базальной мембраной и плазмолеммой симпласта расположены миосателитоциты. Это одноядерные клетки, ядра которых подобные ядер симпласта, но мелкие, круглые и ясные. Клетки имеют органеллы общего призначения, специальные органеллы отсутствуют. Миосателитоциты - это камбиальные элементы волокна, за счет которых происходит процесс роста и регенерации.
Цитоплазма симпласта имеет специальное название - саркоплазма. Ядра, численность которых может достигать нескольких десятка тысяч, как правило, лежат непосредственно под плазмолеммой, имеют удлиненно - овальную фор му, небольшое количество гетерохроматина, в них хорошо заметны ядрышки. В саркоплазме содержатся три группы организованных структуртур: общие органеллы, включенния (жировые, углеводные) и специальные органеллы - миофибриллы. Общие органеллы располагаются, главным образом, у полюсов ядер. Митохондрии крупные, многочисленные, расположены еще и между миофибриллами. Гранулярная эндоплазматическая сеть развита слабо. Агранулярная эндоплазматическая сеть развита очень хорошо, имеет специальное название саркоплазматическая сетка или саркоплазматический ретикулум, особое строение и функцию.
Строение миофибрилл.
Миофибрилы расположенные вдоль мышечного волокна. Длина их совпадает с длиной мышечного волокна, толщина составляет 1-2 мкм. Миофибриллы имеют характерную поперечную исчерченность (чередование светлых и темных полос), что обусловлено особенностью их структуры и в связи с этим различными оптическими свойствами. Вследствие того, что светлые и темные полосы всех миофибрилл отдельного мышечного волокна расположены на одном уровне, все волокно является поперечно - полосатым.
В миофибрилле последовательно расположенные темные анизотропные полосы (или диски А) и светлые изотропные (или диски I). Анизотропные диски окрашиваются интенсивнее, чем изотропные. В поляризованом свете темные полосы имеют двойное променезаломливание - анизотропию, в то время как светлые полосы являются однопроменезаломлеваными (
изотропными) .

This version of Total HTML Converter is unregistered.
Внутри каждой I - полосы является тонкая темная линия, которая имеет название телофрагма, или линии Z. В центре темной А- зоны можно наблюдать более светлый участок - Н - зону, или полоску Гензена, на середине которой находится темная линия М, или мезофрагма. Структурной единицей миофибриллы является саркомер, который представляет собой участок между двумя телофрагмами.
Телофрагма багатая гликозаминогликанами, вследствие чего миофибриллы при мацерации обладают способностью распадатся на отдельные саркомеры (от греческого «саркос» - мясо и «мерос» - часть ). Длина саркомера составляет 2-З мкм.
Структурную формулу саркомера можно записать следующим образом: Т ( Z ) + 1/ 2 І +1 / 2 А + 1/ 2 Н + Н +1 / 2 Н + 1 /2 А + 1/ 2
І + T ( Z ) . Саркомер - это элементарная сократительная единица поперечно - полосатых мышц, который сокращается благодаря тому, что может уменьшать свою длину а два раза.
Под электронным микроскопом в области саркомера были идентифицированы продольные нити, миофиламенты или микрофиламенты, двух типов - тонкие и толстые . Толстые расположенные только в средней части саркомера, построены они из белка миозина.
Тонкие филаменты расположены в I - полосе и частично заходят между толстыми нитями в зону Н. Одним концом они прикрепляются к телофрагме , а другой конец в них свободный, в то время как толстые филаменты имеют оба конца свободных.
Тонкие филаменты построены из белка актина и, кроме того с тропомиозина и тропонина. Диаметр тонких актиновых нитей 5
нм. Толстые миозиновые нити имеют диаметр 10-12 нм и длину 1,5 мкм. Количественное отношение миозиновых нитей к актиновым 1:2 (то есть на один миозиновый миофиламент припадает два актиновых), а взаимное пространственное размещение их гексагональное: на поперечном рассечение тонкие филаменты образуют шестиугольник, в центре которого расположен толстый филамент. Если саркомер в несокращенном состоянии, наиболее темными его частями являются так называемые зоны перекрытия, то есть те части диска А, в которых есть толстые и тонкие миофиламенты. Зона Н выглядит на этом фоне светлой,
так как она состоит только из толстых миозиновых нитей. При сокращении саркомера актиновые филаменты еще дальше проникают в промежуток между миозиновыми, а при полном сокращении их свободные концы почти совпадают внутрь,
которые, очевидно, соединяют срединные участки соседних толстых филаментов. Электронно-микроскопические исследования показали, что Z - линия зигзагообразная, точки прикрепления тонких филаментов на одной стороне Z - пластинки лежат против промежутков между точками прикрепления таких фиментам со второй ее стороны (т.е. соседнего саркомера ). Существует мнение, что Z - пластинка построена из нитей другого типа, так называемых Z - филaмeнтов, которые сочетаются в виде решетки.
Кроме того, Z - линии содержит белок (А - актинин, хотя не установлено, какие именно компоненты Z - линий построены из него.
На электронных микрофотографиях видны короткие нити, которые соединяют между собой актиновые и миозиновые